

Renouées asiatiques envahissantes : la restauration de berges par le génie végétal est-elle si bénéfique à la qualité chimique et à la biodiversité du sol ?
Chapeau
La gestion des renouées asiatiques reste un défi. La non-intervention et les méthodes alternatives telles que la concurrence végétale par génie végétal sont de plus en plus mobilisées alors que peu d’informations sont disponibles sur les conséquences de ces choix de gestion sur l’écosystème, que ce soit sur la composante végétale ou sur le compartiment sol. Cet article présente les résultats d’une étude sur la diversité végétale et le fonctionnement du sol comparant des zones envahies, des zones envahies puis restaurées par concurrence végétale et des zones de référence (i.e. sans invasion).
Contexte de l’étude
Les invasions biologiques posent des enjeux écologiques, économiques et sanitaires à l’échelle mondiale. Parmi les plantes terrestres problématiques en France et en Europe, les renouées asiatiques échappent encore à toute législation. Les renouées asiatiques regroupent la renouée du Japon, Reynoutria japonica Houtt., 1777, la renouée de Sakhaline, Reynoutria sachalinensis (F. Schmidt) Nakai, 1922, et leur hybride nommé la renouée de Bohème, Reynoutria x bohemica Chrtek & Chrtková, 1983. Ces herbacées rhizomateuses vigoureuses se développent en particulier le long des linéaires de transport et des berges de cours d’eau où elles profitent des courants pour se disperser à partir d’un fragment de tige ou de rhizome. Une fois installées, elles forment rapidement des massifs denses et compacts, réduisant fortement la diversité végétale locale et affectant la faune.
Du point de vue des gestionnaires, les renouées asiatiques représentent un véritable enjeu. Le premier choix qui s’offre à eux est de laisser faire. Toutefois, ce choix de non-gestion implique de mieux connaître les conséquences que cela engage, notamment en termes de fonctions écosystémiques. Une autre décision à laquelle sont confrontés les gestionnaires est le choix de la technique de gestion. De plus en plus souvent, du fait de l’efficacité relative des méthodes traditionnelles (fauches répétées, arrachages, traitement chimique, bâchage, etc.), ces derniers essaient des techniques issues du génie végétal comme la replantation de ligneux. Si de nombreuses opérations de génie écologique ont été menées par les gestionnaires, elles n’ont été qu’exceptionnellement suivies dans la durée, et leur efficacité pour restaurer la diversité et le fonctionnement de l’écosystème n’a jamais été évaluée au-delà du suivi des espèces exotiques cibles.
Afin de contribuer à la compréhension des effets des renouées asiatiques sur le compartiment du sol (assemblages biotiques et fonctionnement associé) et de fournir une première évaluation des effets de la restauration par utilisation de la concurrence végétale, nous avons comparé des berges envahies par les renouées asiatiques, des cordons boisés rivulaires non envahis (zones de référence) et des berges envahies puis restaurées par des techniques de génie écologique. Cette comparaison a été menée sur la base de la diversité végétale, des éléments majeurs du sol, des activités enzymatiques de la microflore associées, et de la microfaune et mésofaune endogées. Après dix-huit mois de prospection au sein du bassin versant du Rhône, une sélection de sites a été réalisée. Les sites devaient intégrer des placettes encore envahies par les renouées asiatiques, des placettes restaurées avec replantation de ligneux et des zones non envahies au sein d’un linéaire restreint permettant de supposer une homogénéité de conditions environnementales (photo 1). Sept triptyques de placettes répartis sur deux bassins versants (quatre triptyques le long d’un cours d’eau en Isère – bassin versant 1 ci-après BV1 – et trois triptyques le long d’un cours d’eau dans le département du Rhône – bassin versant 2 ci-après BV2) ont pu être identifiés. Les placettes restaurées correspondent à des berges de cours d’eau ayant été envahies par les renouées asiatiques depuis plusieurs années (trois à plus de vingt ans), puis ayant subi des travaux ciblés pour contenir les massifs de renouées. Elles ont fait l’objet de replantation de végétaux ligneux associée ou non à un semis de mélange grainier herbacé et/ou à un travail préalable du sol, sans paillage ou bâchage. Les placettes de référence non envahies choisies correspondent toutes à des cordons boisés rivulaires (photo 1).
Photo 1 – Exemple de triptyque de placettes au sein d’un site.

Les placettes envahie, restaurée et de référence sont distantes de moins de 1 km les unes des autres au sein de chaque site afin de minimiser les différences de conditions environnementales.
Résultats
Flore
La richesse moyenne en espèces végétales des placettes envahies par les renouées asiatiques est comparable entre bassins versants et varie de onze à seize espèces (figure 1A). Les placettes de référence ont une richesse spécifique autour de vingt-cinq et celle des placettes restaurées ayant fait l’objet de plantations et semis, dépasse les quarante espèces par placette
En effet, si les espèces dominantes des placettes envahies sont bien les renouées asiatiques, la composition végétale n’est pas complètement similaire entre les deux bassins versants échantillonnés. Les espèces les plus fréquentes des placettes envahies et communes aux deux bassins versants sont le lierre terrestre (Glechoma hederacea L., 1753), l’ortie (Urtica dioica L., 1753), le gaillet gratteron (Galium aparine L., 1753), la véronique à feuille de lierre (Veronica hederifolia L., 1753) et la ficaire (Ficaria verna Huds., 1762). Les différences entre les deux bassins versants tiennent surtout aux espèces ligneuses, plus abondantes dans le bassin versant 1. Compte-tenu du blocage de la régénération naturelle lié aux renouées asiatiques, il est probable que les espèces ligneuses présentes aujourd’hui ne puissent plus se régénérer et qu’ainsi les compositions végétales tendent à s’homogénéiser avec le temps.
Figure 1 – Richesse spécifique (A) et indice de diversité de Shannon (B) pour les espèces végétales herbacées et ligneuses des placettes envahies, restaurées et de référence des bassins versants BV1 et BV2.
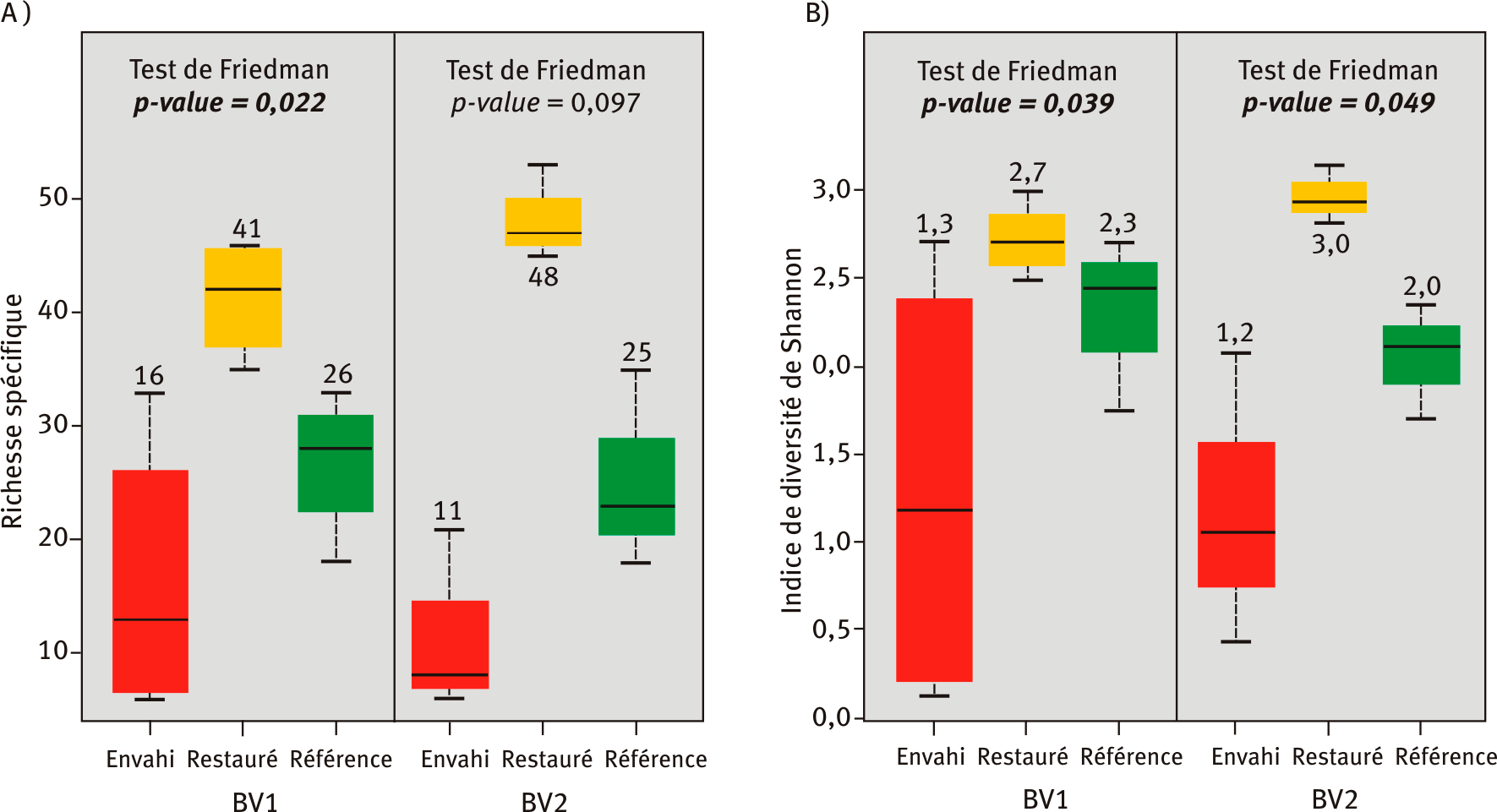
Les placettes envahies, restaurées et de référence des deux bassins versants (BV) étudiés sont symbolisées respectivement en rouge, jaune et vert. Les valeurs indiquées pour les tests de Friedman sont les p-values, calculées au sein de chaque bassin versant. Si la p-value est inférieure à 0,05, alors les trois placettes sont significativement différentes les unes des autres. La p-value est alors indiquée en gras le cas échéant.
Les espèces les plus fréquentes dans les placettes de référence sont le noisetier (Corylus avellana L., 1753), le frêne (Fraxinus excelsior L., 1753), l’érable champêtre (Acer campestre L., 1753), l’aubépine (Crataegus monogyna Jacq., 1775) et la ficaire. Une espèce caractérise spécifiquement les placettes de référence : c’est le lierre grimpant (Hedera helix L., 1753). La ronce (Rubus fruticosus L., 1753) est également très abondante dans le sous-étage. Sur le premier bassin versant, on notera que les ligneux sont plus typiques des cordons rivulaires avec une forte présence d’aulne glutineux (Alnus glutinosa (L.) Gaertn., 1790) et du peuplier noir (Populus nigra L., 1753), tandis que sur le deuxième bassin versant, ce sont des arbres de forêts plus matures qui sont les plus abondants comme le charme (Carpinus betulus L., 1753) ou l’érable sycomore (Acer pseudoplatanus L., 1753).
Les placettes restaurées ont des compositions végétales qui se distinguent nettement de la composition des placettes de référence et des placettes envahies. Cela peut s’expliquer par les espèces choisies par les gestionnaires lors des plantations de restauration qui ne visaient pas spécifiquement à converger vers un milieu rivulaire de référence type (pas de référence explicitement choisie), mais plutôt de recréer une communauté végétale diversifiée dominée par des espèces indigènes, ce qui explique également leur richesse spécifique plus importante. Sur les deux bassins versants, les communautés végétales des placettes restaurées ont des compositions proches, ce qui peut témoigner de pratiques analogues entre les deux organismes de gestion, notamment en termes d’espèces ligneuses utilisées. Ainsi, sur les deux bassins versants, les placettes restaurées hébergent des ligneux comme le frêne, le cornouiller sanguin (Cornus sanguinea L., 1753), l’aubépine, l’érable champêtre, le fusain d’Europe (Euonymus europaeus L., 1753) ou l’églantier (Rosa canina L., 1753), qui sont toutes des espèces plantées dans le cadre de la restauration post-invasion, que l’on retrouve aussi spontanément dans les placettes de référence. Certains végétaux ligneux comme le cornouiller mâle (Cornus mas L., 1753), planté dans les deux bassins versants, le bois puant (Prunus padus L., 1753), le sorbier torminal (Torminalis glaberrima (Gand.) Sennikov & Kurtto, 2017) ou la viorne obier (Viburnum opulus L., 1753) plantés uniquement sur le premier bassin versant, ne sont, en revanche, pas présents de manière spontanée dans les placettes de référence échantillonnées. Du côté des herbacées, certaines espèces présentes dans les placettes envahies et de référence sont aussi très communes dans les placettes restaurées. Il en est ainsi pour le lierre terrestre, la véronique à feuille de lierre, la gaillet gratteron, l’ortie ou la ficaire. D’autres herbacées ont été échantillonnées uniquement dans les placettes restaurées, comme la véronique de Perse (Veronica persica Poir., 1808), la mâche commune (Valerianella locusta (L.) Laterr., 1821), la cardamine (Cardamine hirsuta L., 1753) ou l’anémone fausse renoncule (Anemone ranunculoides L., 1753). Ces herbacées ont été recrutées spontanément sur les placettes restaurées et n’ont pas été recensées dans les placettes de référence, ni dans les placettes envahies. Les renouées asiatiques sont encore présentes, mais leur abondance reste localement faible, en partie grâce à la fauche sélective qui continue d’être appliquée régulièrement sur ces placettes.
Chimie du sol
Éléments majeurs
L’analyse en composante principale (ACP) sur les éléments majeurs du sol (nitrate NO3, ammonium NH4, azote minéral Nminéral, azote total Ntotal, phosphate PO4, phosphore total Ptotal, carbone organique Corg, rapports C/Ntotal, C/Ptotal, N/Ptotal) met en évidence une ségrégation des placettes envahies selon le premier et le second axe respectivement par rapport aux placettes restaurées, plutôt en bas à droite dans le plan de l’ACP (figure 2A). Compte-tenu de leur corrélation avec les différentes variables, les axes 1 et 2 peuvent s’interpréter comme des axes liés à la matière organique et à la disponibilité en nutriments, respectivement. Les placettes restaurées sont ainsi moins riches en matières organiques que les autres types de placettes. Au contraire, les placettes envahies semblent tendre vers un enrichissement en éléments majeurs sans se démarquer toutefois des placettes de référence. Par ailleurs, elles ne convergent pas entre elles, contrairement aux hypothèses de création de niche. En effet, selon cette hypothèse, les renouées asiatiques créeraient des conditions qui leur seraient favorables, stimulant d’autant plus l’enrichissement nutritif dans des sols initialement pauvres, ce qui conduirait à une homogénéisation des conditions nutritives sous leur couvert. Ici, les placettes envahies des deux bassins versants s’écartent nettement les unes des autres, gardant ainsi leur spécificité en termes de composition en éléments majeurs. La largeur des ellipses témoigne par ailleurs de la grande variabilité entre placettes envahies. En revanche, les placettes restaurées présentent une variabilité plus faible ainsi qu’une convergence entre les deux bassins versants. Cela pourrait résulter des travaux de gestion qui ont été réalisés et qui ont pu conduire dans les deux cas à un appauvrissement nutritif du milieu (par exemple dû à un apport exogène de sol minéral et/ou de la suppression de l’horizon supérieur organique lors de l’excavation des renouées asiatiques sur certaines placettes).
Figure 2 – Analyses en composantes principales (ACP) sur les éléments chimiques majeurs (A) et sur les activités enzymatiques (B).
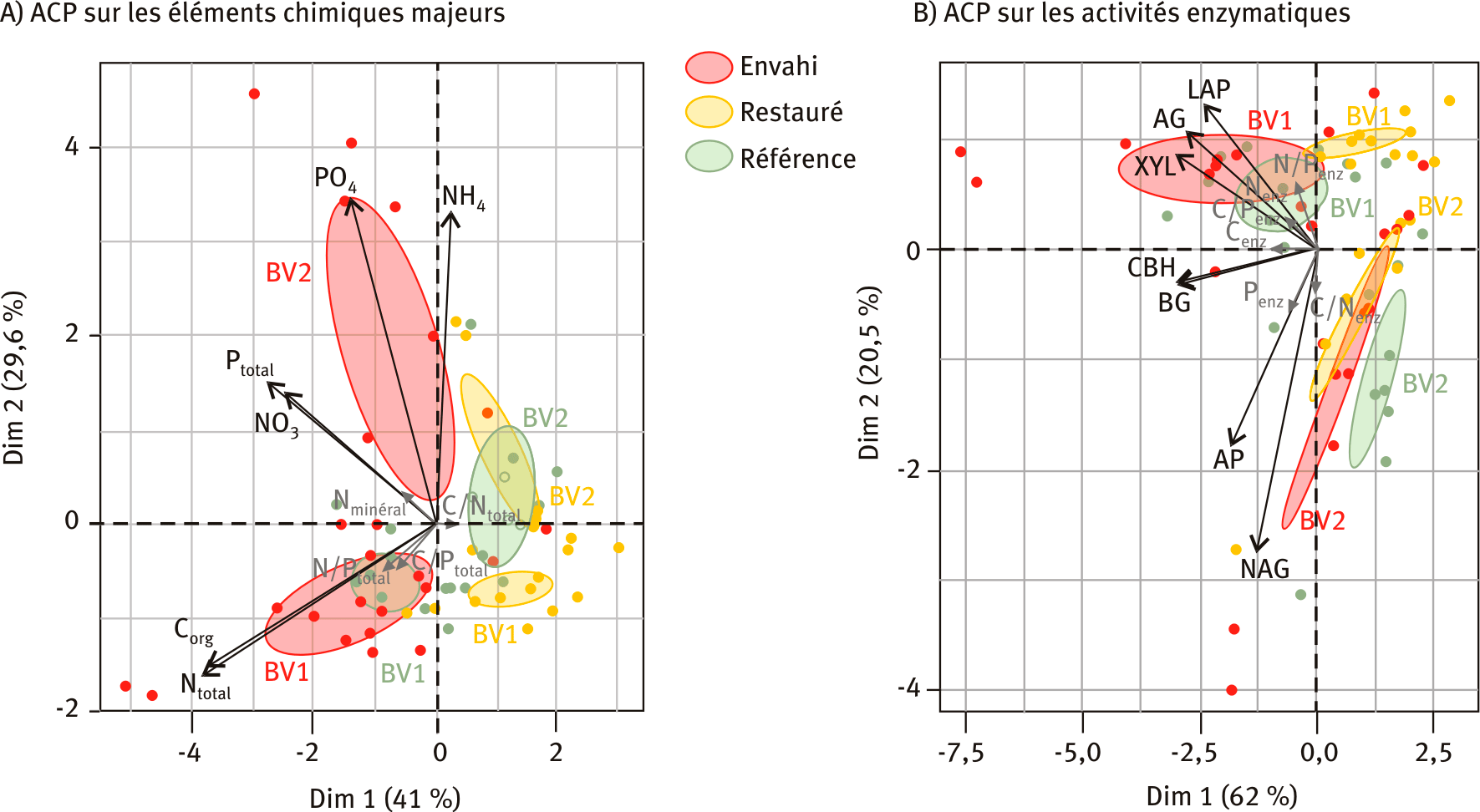
Les placettes envahies, restaurées et de référence des deux bassins versants étudiés (BV1 et BV2) sont symbolisés respectivement en rouge, jaune et vert. Le pourcentage de variabilité représenté sur chaque axe, ou dimension du plan, est indiqué. Les variables projetées en gris sont des variables supplémentaires.
Activités enzymatiques
L’ACP sur les activités enzymatiques liées à la dégradation du carbone organique (β-1,4-glucosidase (BG), α-1,4-glucosidase (AG), 1,4-β-D-cellobiohydrolase (CBH), β-xylosidase (XYL)), à la dégradation de l’azote (β-1,4-N-acetyl-glucosaminidase (NAG), L-leucine aminopeptidase (LAP)) et à la dégradation du phosphore (alkaline phosphatase (AP)) (figure 2B) permet de distinguer nettement les deux bassins versants, et dans une moindre mesure les types de placettes. La position relative des placettes envahies est décalée vers la gauche le long de l’axe 1 pour les deux bassins versants. Cet axe est négativement corrélé aux activités enzymatiques liées au carbone (AG, BG, CBH, XYL), ce qui indique que les activités enzymatiques liées au recyclage du carbone dans le sol sont plus importantes dans les placettes envahies que dans les placettes de référence. Ces résultats ont été confirmés par des tests statistiques (test de Friedman, p-value = 0,0183). Globalement les activités enzymatiques du bassin versant BV2 sont plus faibles que dans le bassin versant BV1. Les placettes restaurées du bassin versant BV1 s’écartent plus nettement des placettes envahies et de référence, présentant une tendance à des activités enzymatiques plus faibles. Les placettes restaurées du BV2 ont une position intermédiaire : les activités enzymatiques y sont légèrement plus faibles que les placettes envahies, mais plus élevées que les placettes de référence.
Microfaune et mésofaune du sol
La densité de nématodes, pour l’ensemble des placettes, est relativement faible suggérant probablement un défaut d’extraction au laboratoire (figure 3A). Néanmoins aucune différence n’apparaît entre les trois types de placettes, même si les placettes restaurées semblent présenter une plus faible densité.
L’abondance totale des acariens ne diffère pas entre les trois types de placettes, quel que soit le bassin versant (figure 3B). Cet ensemble étant constitué d’une grande diversité de groupes fonctionnels ou trophiques, avec des écologies très différentes, leurs réponses à l’invasion peuvent être variées.
L’abondance totale des collemboles diffère entre les trois modalités lorsque les deux bassins versants sont regroupés (p-value = 0,024), avec en moyenne presque trois fois moins d'individus dans les placettes restaurées (3 667 ind.m-2) par rapport aux placettes envahies (10 540 ind.m-2). Cependant, au sein de chaque bassin versant, l’abondance totale de collemboles ne diffère pas entre les modalités, la variabilité étant élevée.
La variabilité de réponse observée dans notre étude peut refléter des différences d’habitat et d’historique de l’occupation du sol ou de gestion. Toutefois, les placettes restaurées montrent un appauvrissement des communautés de collemboles. Au même titre que les résultats obtenus dans la partie sur la chimie du sol, on note un effet négatif de la restauration sur la composante biologique du sol.
Figure 3 – Densité en nématodes (A), abondances en acariens (B) et en collemboles (C) dans les placettes envahies, restaurées et de référence des bassins versants BV1 et BV2.
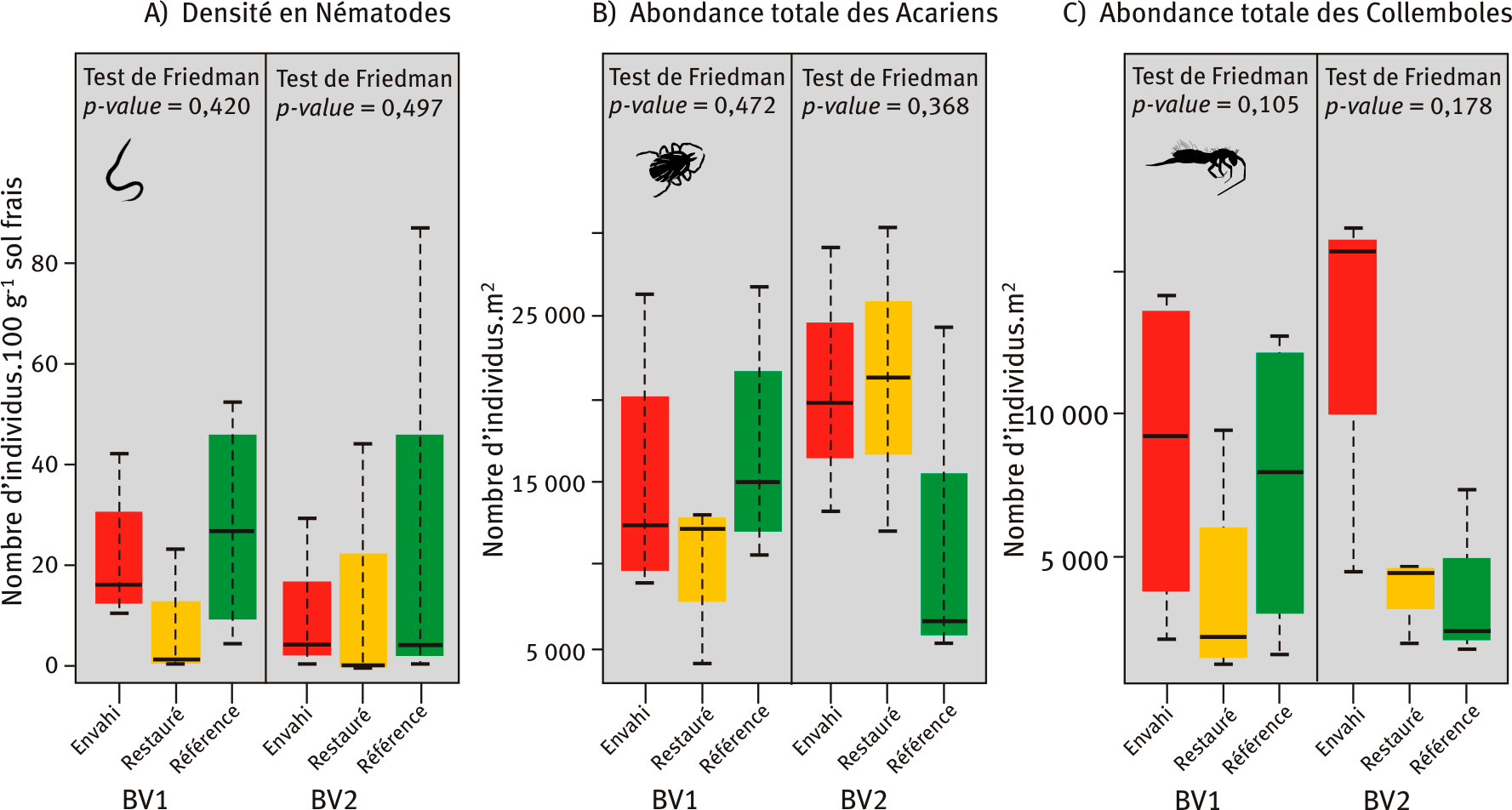
Les placettes envahies, restaurées et de référence des deux bassins versants (BV) étudiés sont symbolisées respectivement en rouge, jaune et vert. Les valeurs indiquées pour les tests de Friedman sont les p-values (ou risque alpha), calculées au sein de chaque bassin versant. Si la p-value est inférieure à 0,05, alors les trois placettes sont significativement différentes les unes des autres. La p-value est alors indiquée en gras le cas échéant.
Conclusions
Des effets négatifs des renouées asiatiques sur la diversité végétale, mais des effets mineurs et variables sur le sol
L’approche observationnelle comparative appliquée sur sept triptyques de placettes réparties sur deux bassins versants montre à la fois des différences entre les placettes envahies et les placettes dites « de référence », mais surtout des différences entre placettes restaurées et les autres placettes. Ainsi les renouées asiatiques ont eu tendance à enrichir le sol en matière organique et à augmenter la disponibilité en nutriments et les activités enzymatiques du sol en comparaison des placettes de référence (encadré 1). La biologie du sol est également peu affectée, avec une tendance à la stimulation des communautés de collemboles dont les abondances sont plus importantes dans les placettes envahies. Par ailleurs, le faible effet mesuré sur les variables du sol en réponse à l’envahissement par les renouées asiatiques peut s’expliquer par une forte variabilité entre les sites, et particulièrement au sein des placettes envahies. Cette variabilité va à l’encontre de l’hypothèse de création de niche et d’homogénéisation du milieu lors de l’invasion par les renouées asiatiques. Une plus large étude comparant magnitude et direction des effets des renouées et conditions initiales permettrait d’évaluer si leurs effets sur les concentrations en éléments majeurs du sol dépendent de la richesse initiale du milieu et si la présence de renouées asiatiques conduit ainsi à la modification du niveau nutritif du sol.
En revanche, nos résultats montrent clairement que les renouées asiatiques appauvrissent la richesse taxonomique et la diversité des espèces végétales comparativement aux écosystèmes de référence choisis. En colonisant le milieu de manière dominante, elles excluent les autres espèces par compétition directe, notamment pour la ressource lumineuse, ou indirecte par sécrétions de composés allélopathiques. Toutefois, certaines espèces de plantes sont capables de coexister avec les renouées asiatiques, et notamment les espèces précoces comme la ficaire ou le gaillet gratteron.
La restauration est positive pour la diversité végétale mais impacte négativement le sol, plus que l’invasion elle-même
Sur le sol, les effets de la restauration sont plus nets que les effets des renouées asiatiques qui apparaissent comme peu significatifs et hautement variables. Ainsi, un appauvrissement nutritif en éléments majeurs est observé sur les placettes restaurées des deux bassins versants étudiés. On observe également dans une moindre mesure de plus faibles activités enzymatiques et un appauvrissement de la mésofaune du sol. Pourtant, la diversité végétale est beaucoup plus importante dans les placettes restaurées que dans les placettes envahies, ainsi que dans les placettes de référence. La restauration entraîne donc une déconnection des compartiments épigé (flore) et endogé (sol). Ce résultat est étonnant car les pratiques de gestion étudiées sont réputées avoir peu d’impact sur le milieu comparativement à d’autres méthodes comme le broyage-concassage, par exemple, qui nécessite une extraction complète de la biomasse souterraine et un remaniement profond du sol. Toutefois, les placettes restaurées sélectionnées ont parfois fait l’objet d’un travail préalable du sol, plus ou moins important, allant du griffage de surface à l’importation de terres et au remodelage de la berge, qui pourrait expliquer que le fonctionnement du sol ait été perturbé. Ainsi, il parait important de mieux évaluer dans de futures études quelles sont les méthodes de restauration mises en place au niveau des sols, et notamment s’il existe des méthodes permettant de diminuer les impacts négatifs d’un retournement profond, de la suppression de l’horizon organique supérieur ou d’un apport exogène de sol minéral pauvre en nutriments. Bien que des méthodes de restauration du sol restent à étudier, les placettes restaurées n’ont montré qu’une faible corrélation entre leur écart à la référence et l’intensité de travail du sol dont elles ont fait l’objet, ce qui laisse penser que d’autres facteurs (ex. : histoire des sites, méthodes de restauration, pédoclimat) peuvent également influencer le fonctionnement du sol. En revanche, le temps semble effacer les écarts : les placettes restaurées depuis plus d’une dizaine d’années présentaient un fonctionnement qui se rapprochait plus de celui des placettes de référence que des parcelles restaurées récemment (encadré 2). En d’autres termes, l’impact négatif de la restauration sur le fonctionnement du sol semble se résorber au bout d’une dizaine d’année.
Ces résultats, s’ils restent limités à l’exemple des deux bassins versants étudiés, soulignent que l’effort de restauration sur la diversité végétale porte ses fruits. Cependant, le compartiment du sol ne doit pas être négligé et les impacts anthropiques sur son fonctionnement, y compris lorsqu’il s’agit d’avoir une action positive sur le milieu comme la restauration écologique post-envahissement, devraient être intégrés dès l’amont du projet de gestion.
Encadré 1 – Les effets des renouées asiatiques sur le sol sont variables et contexte-dépendants, éclairage de la bibliographie scientifique.
De nombreuses publications s’intéressent aux impacts des renouées asiatiques sur le sol, avec un focus ces dernières années sur son fonctionnement biologique. Les conclusions de ces études sont très variables, allant d’effets significatifs à une absence d’effets, et mettant en évidence la contexte-dépendance de l’effet des renouées asiatiques sur le sol.
Les résultats de notre étude sont ainsi cohérents avec la majorité des publications testant les différences d’azote total et minéral entre sol envahi par des renouées asiatiques et milieu de référence. Dans la plupart des cas, elles ne trouvent aucun effet 3-10, sauf pour le nitrate où certaines études mettent en évidence un enrichissement lié à l’invasion 6,8,11-13 et deux cas où l’invasion a un effet négatif sur l’azote 12,14. Les résultats de la littérature concernant le carbone organique sont aussi variables et la plupart rendent compte d’effets négligeables 3,7-9,14, même si une publication rend compte d’effets positifs des renouées asiatiques 10. Concernant le phosphore et le phosphate, les résultats de la littérature scientifique sont tout aussi variables, des articles attestant d’effets négligeables 5,9,14, d’autres d’effets négatifs 12,13.
Les activités enzymatiques (qui reflètent l’activité biologique du sol) sous les renouées asiatiques peuvent être modifiées dans un sens positif ou négatif selon la profondeur 15 ou les enzymes considérées 16 ou être similaires à la référence 17,18.
Nos résultats montrent que la présence de renouées asiatiques semble peu affecter les abondances totales de microarthropodes ou microfaune du sol 7,19. Toutefois, à l’inverse de certaines études 11, nous montrons un effet plutôt positif des renouées asiatiques sur les assemblages de collemboles. D’autres 11,20 pointent dans leurs études l’importance de l’habitat (type, structure) envahi quant à l’impact des renouées sur les communautés d’organismes du sol.
Cette variabilité continue d’interroger et d’intéresser la recherche. Si la variabilité temporelle des effets des renouées asiatiques sur l’azote à l’échelle d’une saison de végétation a été mise en lien avec leur cycle biologique 21, la variabilité spatiale de leurs effets semble liée au type de sol 3,10,18,22,23, au type d’écosystème de référence 11,24,25 ou à la profondeur d’échantillonnage 15,21. Par ailleurs, leurs effets sur la biologie du sol semblent dépendre de la dose de composés secondaires 10,26 et encore une fois du type d’écosystème comparé 11. Les déterminants de cette variabilité et les mécanismes sous-jacents restent donc à explorer pour tenter de généraliser et prédire les effets des renouées asiatiques.
_______________________________________________________________________________
1. Lavoie, C. (2017).The impact of invasive knotweed species (Reynoutria spp.) on the environment: review and research perspectives. Biol Invasions, 19, 2319-2337. https://doi.org/10.1007/s10530-017-1444-y
2. Dommanget, F., Evette, A. & Piola, F. (2019). État de l’art des techniques de génie végétal pour contrôler les renouées. Sciences Eaux & Territoires, 27, 74-79. https://doi.org/10.14758/SET-REVUE.2019.1.13
3. Dassonville, N., Guillaumaud, N., Piola, F., Meerts, P. & Poly, F. (2011). Niche construction by the invasive Asian knotweeds (species complex Fallopia): impact on activity, abundance and community structure of denitrifiers and nitrifiers. Biol Invasions, 13, 1115-1133. https://doi.org/10.1007/s10530-011-9954-5
4. Aguilera, A. G., Alpert, P., Dukes, J. S. & Harrington, R. (2010). Impacts of the invasive plant Fallopia japonica (Houtt.) on plant communities and ecosystem processes. Biol. Invasions, 12, 1243-1252. https://doi.org/10.1007/s10530-009-9543-z
5. Dassonville, N., Vanderhoeven, S., Gruber, W. & Meerts, P. (2007). Invasion by Fallopia japonica increases topsoil mineral nutrient concentrations. Ecoscience, 14, 230-240. https://doi.org/10.2980/1195-6860(2007)14[230:IBFJIT]2.0.CO;2
6. Dommanget, F., Evette, A., Spiegelberger, T., Gallet, C., Pacé, M., Imbert, M. & Navas, M. L. (2014). Differential allelopathic effects of Japanese knotweed on willow and cottonwood cuttings used in riverbank restoration techniques. Journal of Environmental Management, 132, 71-78. https://doi.org/10.1016/j.jenvman.2013.10.024
7. Schmitz, D., Girardi, J., Ullrich, E., Muñoz-Sepulveda, K., Bundschuh, M., Riess, K. & Schirmel, J. (2023). Fallopia japonica and Impatiens glandulifera are colonized by species-poor root-associated fungal communities but have minor impacts on soil properties in riparian habitats. Biological Invasions, 25, 2199-2218. https://doi.org/10.1007/s10530-023-03034-2
8. Stefanowicz, A. M., Stanek, M., Nobis, M. & Zubek, S. (2017). Few effects of invasive plants Reynoutria japonica, Rudbeckia laciniata and Solidago gigantea on soil physical and chemical properties. Science of The Total Environment, 574, 938-946. https://doi.org/10.1016/j.scitotenv.2016.09.120
9. Woch, M. W., Kapusta, P., Stanek, M., Zubek, S. & Stefanowicz, A. M. (2021). Functional traits predict resident plant response to Reynoutria japonica invasion in riparian and fallow communities in southern Poland. AoB PLANTS, 13 (4), plab035. https://doi.org/10.1093/aobpla/plab035
10. Zhang, Z. & Suseela, V. (2021). Nitrogen availability modulates the impacts of plant invasion on the chemical composition of soil organic matter. Soil Biology and Biochemistry, 156, 108195. https://doi.org/10.1016/j.soilbio.2021.108195
11. De Almeida, T., Forey, E. & Chauvat, M. (2022). Alien invasive plant effect on soil fauna is habitat dependent. Diversity, 14(2), 61. https://doi.org/10.3390/d14020061
12. Stefanowicz, A. M., Majewska, M. L., Stanek, M., Nobis, M. & Zubek, S. (2018). Differential influence of four invasive plant species on soil physicochemical properties in a pot experiment. J Soils Sediments, 18, 1409-1423. https://doi.org/10.1007/s11368-017-1873-3
13. Zubek, S., Majewska, M. L., Błaszkowski, J., Stefanowicz, A. M., Nobis, M. & Kapusta, P. (2016). Invasive plants affect arbuscular mycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biology and Fertility of Soils, 52, 879-893. https://doi.org/10.1007/s00374-016-1127-3
14. Stefanowicz, A. M., Banaś, A., Stanek, M., Woch, M. W. & Zubek, S. (2020). Large differences in biomass quantity and quality between invasive Reynoutria japonica and resident vegetation are not reflected in topsoil physicochemical properties. Geoderma, 368, 114307. https://doi.org/10.1016/j.geoderma.2020.114307
15. Min, K. & Suseela, V. (2020). Plant invasion alters the Michaelis–Menten kinetics of microbial extracellular enzymes and soil organic matter chemistry along soil depth. Biogeochemistry, 150, 181-196. https://doi.org/10.1007/s10533-020-00692-5
16. Suseela, V., Alpert, P., Nakatsu, C. H., Armstrong, A. & Tharayil, N. (2016). Plant–soil interactions regulate the identity of soil carbon in invaded ecosystems: implication for legacy effects. Functional Ecology, 30, 1227-1238. https://doi.org/10.1111/1365-2435.12591
17. Stefanowicz, A. M., Frąc, M., Oszust, K. & Stanek, M. (2022). Contrasting effects of extracts from invasive Reynoutria japonica on soil microbial biomass, activity, and community structure. Biological Invasions, 24, 3233-3247. https://doi.org/10.1007/s10530-022-02842-2
18. Stefanowicz, A. M., Kapusta, P., Stanek, M., Frqc, M., Oszust, K., Woch, M. W., Zubek, S. (2021). Invasive plant Reynoutria japonica produces large amounts of phenolic compounds and reduces the biomass but not activity of soil microbial communities. Science of The Total Environment, 767, 145439. https://doi.org/10.1016/j.scitotenv.2021.145439
19. Skubała, P. & Mierny, A. (2009). Invasive Reynoutria taxa as a contaminant of soil. Does it reduce abundance and diversity of microarthropods and damage soil habitat? Pesticides 1, 57-62.
20. Abgrall, C., Forey, E. & Chauvat, M. (2019). Soil fauna responses to invasive alien plants are determined by trophic groups and habitat structure: a global meta‐analysis. Oikos, 128, 1390-1401. https://doi.org/10.1111/oik.06493
21. Tharayil, N., Alpert, P., Bhowmik, P. & Gerard, P. (2013). Phenolic inputs by invasive species could impart seasonal variations in nitrogen pools in the introduced soils: A case study with Polygonum cuspidatum. Soil Biology and Biochemistry, 57, 858-867. https://doi.org/10.1016/j.soilbio.2012.09.016
22. Koutika, L.-S., Vanderhoeven, S., Chapuis-Lardy, L., Dassonville, N. & Meerts, P. (2007). Assessment of changes in soil organic matter after invasion by exotic plant species. Biology and Fertility of Soils, 44, 331-341. https://doi.org/10.1007/s00374-007-0210-1
23. Vanderhoeven, S., Dassonville, N. & Meerts, P. (2005). Increased topsoil mineral nutrient concentrations under exotic invasive plants in Belgium. Plant Soil, 275, 169-179. https://doi.org/10.1007/s11104-005-1257-0
24. Lecerf, A., Patfield, D., Boiché, A., Riipinen, M. P., Chauvet, E. & Dobson, M. (2007).Stream ecosystems respond to riparian invasion by Japanese knotweed ( Fallopia japonica ). Can. J. Fish. Aquat. Sci., 64, 1273-1283. https://doi.org/10.1139/f07-092
25. Stefanowicz, A. M., Stanek, M., Majewska, M. L., Nobis, M. & Zubek, S. (2019). Invasive plant species identity affects soil microbial communities in a mesocosm experiment. Applied Soil Ecology, 136, 168-177. https://doi.org/10.1016/j.apsoil.2019.01.004
26. Abgrall, C., Forey, E., Mignot, L. & Chauvat, M. (2018). Invasion by Fallopia japonica alters soil food webs through secondary metabolites. Soil Biology and Biochemistry, 127, 100-109. https://doi.org/10.1016/j.soilbio.2018.09.016
Encadré 2 – Les effets négatifs de la restauration sur le sol se résorbent avec le temps.
Afin de regarder comment évolue l’écart des placettes restaurées par rapport aux placettes de référence, nous avons calculé le « log-response-ratio » (LRRMoy), soit le logarithme népérien du rapport entre les valeurs pour les placettes restaurées et la valeur moyenne des placettes de référence de chaque bassin versant, sur chaque élément majeur synthétique du sol (carbone, azote total et phosphore total) et sur les activités enzymatiques associées. Cet indice permet de comparer les différences entre les placettes restaurées et la référence moyenne du bassin versant. Si le LRRMoy est négatif, alors les placettes restaurées ont des valeurs inférieures aux placettes de référence ; si le LRRMoy est nul, alors les placettes restaurées n’ont pas de différences avec la référence et si le LRR est positif, alors les placettes restaurées ont des valeurs supérieures à la référence.
Figure 4
Les résultats montrent que le LRRMoy est significativement corrélé avec l’âge de l’ouvrage, que ce soit pour les éléments majeurs testés ou dans une moindre mesure les activités enzymatiques. Ainsi, l’écart des placettes à la référence s’atténue au cours du temps et au bout d’une douzaine d’années, semble se réduire complètement. L’appauvrissement en éléments majeurs du sol et la plus faible activité enzymatique détectés sur les placettes restaurées tend à se résorber, mais il semblerait qu’une dizaine d’années soit nécessaire pour atteindre des valeurs comparables à celles des références.
_______________________________________
Photo d’entête : © Tomas Vynikal – Adobe Stock
Notes
- « Restauration » est ici entendu au sens large et s’appuie sur la définition proposée par la Society for Ecological Restoration : la restauration écologique est un processus qui aide au rétablissement d'un écosystème dégradé, endommagé ou détruit (Gann et al., 2019, International principles and standards for the practice of ecological restoration, Restoration Ecology, 27 (51), S1-S46).
- L’indice de Shannon prend en compte le nombre d’espèces d’une communauté et la répartition des individus au sein de ces espèces, permettant d’en quantifier l’hétérogénéité. Ainsi, un faible indice de Shannon peut exprimer un faible nombre d’espèces et/ou des individus répartis de manière très hétérogène au sein de ces espèces et inversement, un indice de Shannon élevé peut exprimer un grand nombre de d’espèces avec des abondances relatives homogènes.
Références
- Dommanget, F., Evette, A., & Piola, F. (2019). État de l’art des techniques de génie végétal pour contrôler les renouées. Sciences Eaux & Territoires, 27, 74-79. https://doi.org/10.14758/SET-REVUE.2019.1.13
- Dommanget, F., & Duc, H. (2019). Portrait esquissé du génie végétal en tant qu’outil de gestion des renouées asiatiques. Lettre d’Information Centre de Ressources EEE n° 6, http://especes-exotiques-envahissantes.fr/portrait-esquisse-du-genie-vegetal-en-tant-quoutil-de-gestion-des-renouees-asiatiques/
- De Almeida, T., Forey, E., & Chauvat, M. (2022). Alien invasive plant effect on soil fauna is habitat dependent. Diversity,14, 61. https://doi.org/10.3390/d14020061
Résumé
Les espèces exotiques envahissantes représentent une menace pour la biodiversité à l’échelle mondiale. Parmi les plantes problématiques en France et en Europe, les renouées asiatiques (Reynoutria spp.) posent un véritable défi, leur gestion étant coûteuse et d’efficacité aléatoire. Premièrement, les gestionnaires doivent choisir d’agir ou de ne pas intervenir. Pour décider, il leur faut mieux connaître les conséquences (y compris de la non-intervention) sur le fonctionnement de l’écosystème. Ensuite, ils doivent décider de la technique de gestion. Les techniques issues du génie végétal basées sur la réintroduction d’espèces végétales indigènes compétitives permettent de réduire la dominance de l’espèce exotique envahissante tout en réintroduisant de la diversité végétale. Si de nombreuses opérations de génie écologique ont été menées, leur efficacité pour restaurer le fonctionnement de l’écosystème n’a jamais été évaluée, encore moins sur la diversité et l’activité des organismes du sol. Cet article synthétise les résultats d’une étude portant sur les effets des renouées asiatiques et de la restauration des berges envahies par génie végétal sur la communauté végétale et le sol. Il s’appuie notamment sur une analyse fine de la chimie et de la faune du sol dans deux bassins versants (en Isère et dans le Rhône).
Pas de document complémentaire pour cet article
Statistiques de l'article
 Vues: 9284
Vues: 9284
Téléchargements
 PDF: 431
PDF: 431
 XML: 65
XML: 65