

Construction d’un modèle d’estimation du nombre de lamproies sur frayère en utilisant un simple comptage de nids
La lamproie marine est un poisson migrateur en danger d’extinction, notamment en France. Afin d’évaluer l’effectif de reproducteurs directement sur les frayères, un modèle individu-centré utilisant un comptage de nids a été développé et intégré dans une application simple d’utilisation. Les résultats obtenus sont proches de ceux d’un modèle de capture-marquage-recapture classique et cet outil peut aisément être adapté en fonction des besoins des gestionnaires.
Introduction
L’estimation de l’effectif d’une population nécessite de prendre en compte la biologie de l’espèce considérée, mais aussi les contraintes de faisabilité, de coût et de nombre de personnes impliquées. Une première approche est le comptage direct des individus afin d’estimer un effectif. Une méthode classique est la capture, le marquage puis la recapture (CMR) des individus, qui présente pour avantage de fournir une taille de population assez précise (Funk et al., 2003) tout en étant aisément adaptable aux particularités de l’espèce considérée (photo 1). Ainsi, il est possible de prendre en compte l’entrée ou non d’individus sur le site d’étude (Schwarz et Seber, 1999) ou encore de ne pas recapturer les individus après la capture initiale (McClintock et White, 2009). Malgré son adaptabilité, cette méthode nécessite de capturer une part importante de la population, et implique donc un grand effort d’échantillonnage induisant des coûts logistiques et financiers souvent trop importants pour les structures en charge des programmes de suivi.
Photo 1. Capture d’individus sur un nid lors du protocole de capture-marquage-recapture visant à calibrer le modèle.

Une seconde approche est l’utilisation de méthodes basées sur des indices de présence d’une espèce et non sur un comptage direct d’individus. Cette approche peut notamment limiter les problèmes de faible détectabilité ou de marquage. Parmi les indices fréquemment employés, on retrouve les barrages de castors (Johnston et Windels, 2015) ou encore les nombreux indices de présence d’oiseaux, parmi lesquels les chants, les fèces ou encore les nids (Morgan et al., 1983). Cependant, ces méthodes ne produisent généralement pas d’estimation absolue de population mais une estimation relative. Pour obtenir une estimation absolue, c’est-à-dire une estimation d’un nombre d’individus et non d’un nombre d’indices, il est nécessaire d’établir la relation entre l’indice et l’espèce considérée, par exemple via une régression. Cela peut être fait en collectant à la fois le nombre d’indices et le nombre d’individus sur de multiples sites. Néanmoins, cette relation dépend de multiples paramètres parmi lesquels les conditions environnementales ou les caractéristiques individuelles. Pour prendre en compte ces différents éléments de variabilité, un nombre important d’observations est souvent nécessaire, ce qui est en général coûteux, voire impossible à obtenir.
Parmi les espèces dont l’estimation des populations présente un intérêt particulier, on retrouve la lamproie marine, Petromyzon marinus, qui est un poisson migrateur anadrome
– collecte d’informations sur le comportement menant à la construction des nids et prise en compte de la variabilité individuelle (notamment la polygamie, la construction de plusieurs nids par un même individu et la variabilité de date d’arrivée sur la frayère) ;
– construction d’un modèle simulant la construction de nids à partir d’un nombre donné d’individus ayant chacun un ensemble de paramètres basé sur les informations collectées ;
– comparaison du nombre de nids simulé par le modèle au nombre de nids observés sur le terrain pour obtenir une estimation absolue de la population avec son incertitude.
Le modèle employé est un modèle bayésien de type ABC
La suite de l’article décrit le fonctionnement du modèle mis en œuvre et présente les estimations obtenues, mais aussi les limites et pistes d’amélioration envisagées pour une utilisation par les gestionnaires intéressés par l’estimation des effectifs de lamproie marine.
Le modèle développé (Dhamelincourt et al., 2023) est mécaniste individu-centré et de type ABC, utilisant l’algorithme de Lenormand (Lenormand et al., 2013). En pratique, il simule une saison de reproduction de lamproie marine avec un pas de temps journalier, pour un pool d’individus mâles et femelles donné (figure 1). Chaque individu possède des caractéristiques déterminées par les paramètres du modèle et estimées d’après des observations menées sur la Nive durant la saison de reproduction de 2019 (Dhamelincourt et al., 2021a), sur un bras court-circuité par la centrale hydro-électrique d'Halsou au Pays Basque (coordonnées : 43° 22’ 21.38” N, 1° 24’ 53.134” W).
Figure 1. Fonctionnement du modèle ABC adapté à la simulation d’une période de reproduction de lamproie marine.
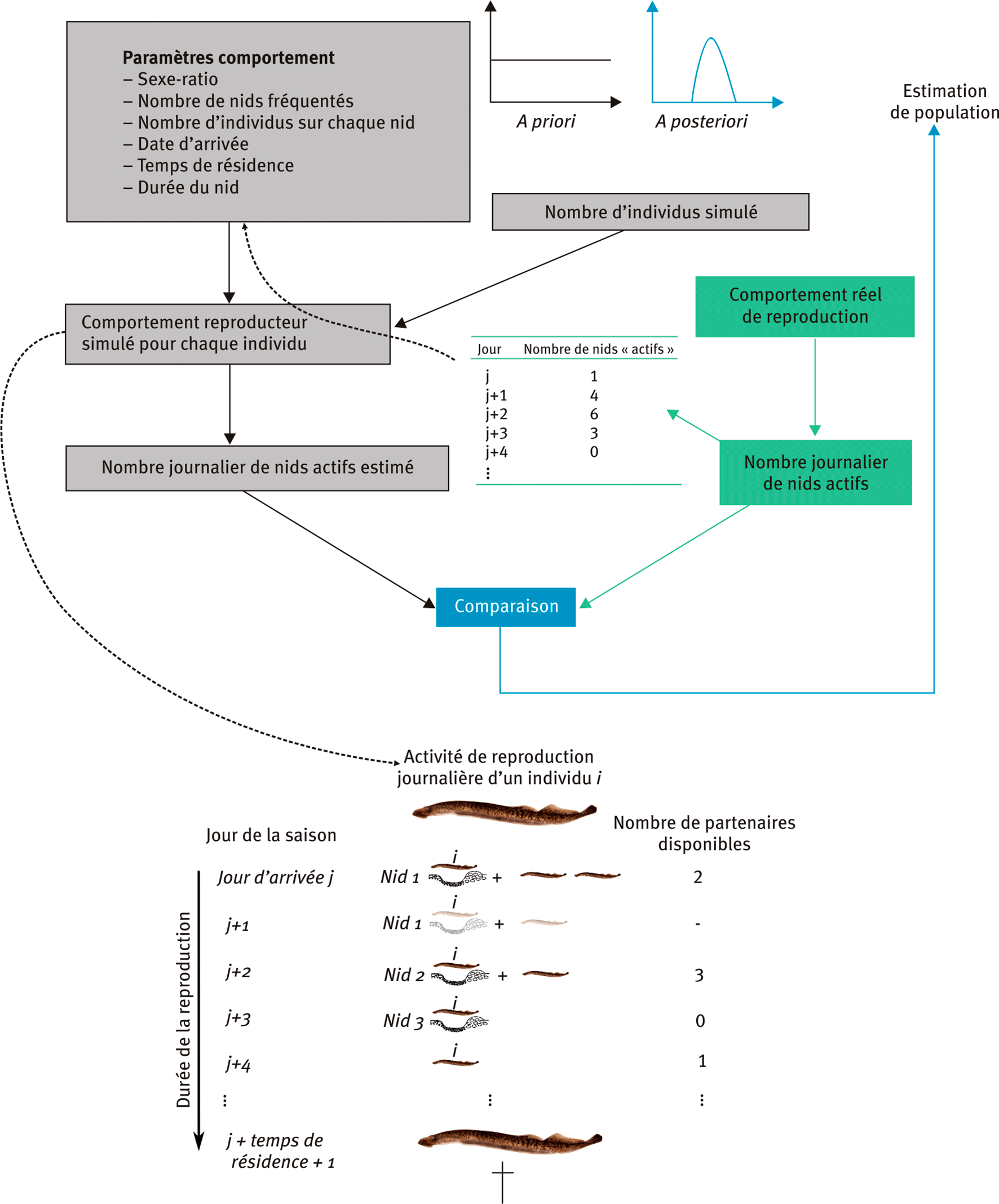
Chaque jour j de la saison de reproduction, un individu peut participer à la construction d’un nid selon les paramètres propres à cet individu, selon son historique de construction durant les jours 1 à j-1 mais également selon les paramètres et l’historique des individus avec lesquels il est amené à interagir (induisant la disponibilité ou non de ces individus).
Chaque individu possède un ensemble de paramètres dépendant du sexe, qui sont le nombre de nids fréquentés, le jour d’arrivée sur la frayère et le temps de résidence. Dans le détail, pour un jour j donné de la saison de reproduction, un individu peut construire un nid avec d’autres individus si cet individu et ses partenaires sont considérés comme actifs (c’est-à-dire que le jour considéré est compris entre la date d’arrivée sur la frayère et cette date + le temps de résidence). Ensuite, cet individu ne peut pas construire plus d’un nombre donné de nids. L’individu est impliqué dans la construction d’un seul nid à la fois. Un nid ne peut pas être construit par plus de mâles et de femelles qu’une valeur limite, aléatoire pour chaque nid et dépendante des observations de 2019. Dans le cas où un individu est actif mais ne trouve pas de partenaire, il peut initier la construction d’un nid sans être rejoint, comme cela a été observé sur le terrain, à la fois pour les mâles et les femelles (cependant plus rarement pour ces dernières).
Ce processus est répété chaque jour jusqu’à la fin de la saison de reproduction, quand tous les individus ont atteint leur nombre maximal de nids et/ou leur temps de résidence. Le modèle fournit alors un nombre journalier de nids actifs (nids sur lesquels au moins un individu creuse un jour donné).
Après k simulations de saisons de reproduction (k dépendant de l’algorithme de Lenormand utilisé, qui détermine automatiquement le nombre de simulations), chacune réalisée avec un nombre Nk d’individus différent, le modèle fournit une distribution du nombre d’individu Nestimate le plus vraisemblable selon les statistiques résumant le jeu de données réellement observé. Les statistiques employées ici sont le nombre maximal, la médiane, la moyenne, le quantile à 25 % et le quantile à 75 % du nombre de nids construits sur une journée.
Pour plus de détails sur le fonctionnement du modèle ainsi que sur les distributions des paramètres, consulter Dhamelincourt et al. (2023).
Le comptage des nids actifs pendant 28 jours entre le 9 mai et le 24 juin 2019 a permis de détecter 146 nids actifs (par jour : 0 nid au minimum et 15 nids au maximum). À partir de ces données, le modèle ABC estime un effectif de 148 individus, avec un intervalle de confiance à 95 % entre 115 et 182 individus. L'effectif estimé est 25 % inférieur à celui estimé par un modèle de CMR (197 individus avec un intervalle de confiance à 95 % situé entre 168 et 236 individus) mis en œuvre lors de la même saison de reproduction (voir figure 2 pour les distributions des deux types de modèles). Le modèle de CMR est un modèle d’occupation multi-états dynamique avec population augmentée (pour plus de détails, voir Dhamelincourt et al., 2021a ; Kery et Schaub, 2011). Cette différence significative d’estimation (d’après un test statistique de Wilcoxon
– des paramètres journaliers comme la densité d’individus peuvent influencer leur répartition dans les nids par le biais d’une modification des paramètres individuels, définis à l’échelle de la saison de reproduction. Par exemple, nous avons pu observer des pics d’activité allant jusqu’à 15 nids actifs sur une journée. Or ce genre de configuration est susceptible d’augmenter la coopération pour la construction des nids, et donc le nombre d’individus par nid ;
– les paramètres du modèle influençant le plus le nombre d’individus estimé peuvent manquer de précision dans leur estimation. Une analyse de sensibilité réalisée dans Dhamelincourt et al. (2023) a identifié la date d’arrivée, le sex-ratio, le nombre de nids par individu et la durée d’activité comme étant les paramètres modifiant le plus l’estimation finale.

Ces résultats montrent donc la nécessité de compléter les estimations des paramètres implémentés dans le modèle, en particulier ceux influençant le plus les estimations. La variabilité biologique des paramètres est prise en compte via leur variance, hormis pour le sex-ratio qui est fixé à 0,5 (autant de mâles que de femelles observés en 2019). Cependant, ils sont pour le moment estimés sur un seul site d’une seule rivière, la Nive à Halsou. Il apparaît donc nécessaire de déterminer leurs valeurs et leur variabilité sur d’autres cours d’eau, de caractéristiques hétérogènes. Cela permettrait notamment de quantifier la variabilité des paramètres entre les cours d’eau et de comprendre quels sont les facteurs affectant cette variabilité.
Par ailleurs, la comparaison entre le modèle de CMR et le modèle ABC pourrait être répétée sur ces mêmes cours d’eau pour évaluer l’écart d’estimation entre les deux méthodes.
Utilisation du modèle par les gestionnaires
Afin de permettre une utilisation simple par les gestionnaires intéressés par l’estimation des populations de lamproie marine, sans avoir à utiliser le code R
Cette interface comprend cinq onglets :
1) Description de l’application : cet onglet informe l’utilisateur sur le but de l’application et sur comment l’utiliser.
2) Chargement des données et du nombre d’individus a priori : cet onglet permet à l’utilisateur de charger ses données. Le fichier de données doit être au format .csv et comporter une première colonne indiquant le jour de suivi (une ligne par jour de suivi) et une seconde colonne indiquant le nombre de nids actifs comptés ce jour-là. Un nid actif est un nid sur lequel au moins un individu est observé le jour considéré, indépendamment de si ce nid était présent ou occupé les jours précédents. L'utilisateur peut indiquer le type de séparateur et la présence d'en-têtes, et visualiser le tableau entré. C'est également sur cette page que l'utilisateur indique les bornes inférieure et supérieure du nombre d'individus sur la zone a priori, selon son expérience ou des données préliminaires. Cette valeur permet d’optimiser le temps de calcul en permettant au modèle de tester des valeurs de population réalistes pour le site considéré.
3) Visualisation et altération des paramètres : la distribution statistique de chaque paramètre du modèle est montrée sous forme d'histogramme. Si l'utilisateur dispose de données lui indiquant que d'autres valeurs s'appliquent à sa population d'intérêt, il peut modifier la moyenne et la dispersion de chaque distribution. Les graphiques changent automatiquement quand les valeurs sont modifiées. Les simulations utiliseront alors ces nouvelles distributions.
4) Lancement de l’analyse et visualisation des résultats : lorsque les données sont chargées et que les paramètres sont vérifiés, l’utilisateur peut lancer l’analyse en cliquant sur le bouton correspondant. Après quelques minutes (temps dépendant notamment du jeu de données), un graphique indiquant la distribution a posteriori du nombre de lamproies apparaît, sur lequel sont indiqués la médiane et les bornes de l’intervalle à 95 %. Un tableau indique également ces valeurs.
5) Contacts et références : cet onglet donne accès aux articles liés au projet, aux contacts des développeurs ainsi qu’aux sites web des partenaires.
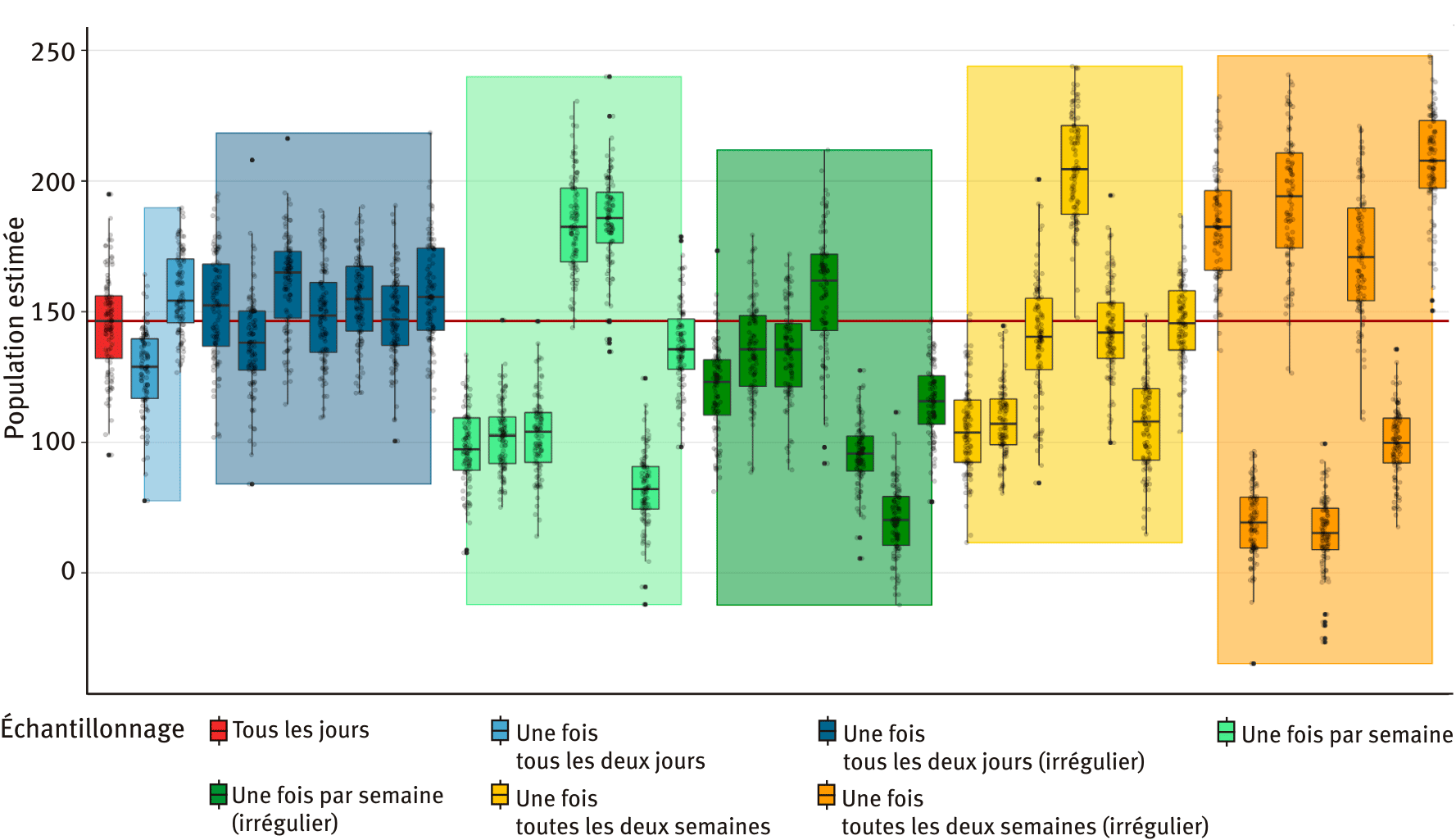
L’objectif du modèle étant d’être utilisé pour le suivi des populations par les gestionnaires, il apparaît nécessaire de tester différentes fréquences d’échantillonnage afin de limiter le temps passé sur le terrain tout en conservant des résultats exploitables. La sensibilité du modèle au protocole d'échantillonnage a donc été testée en faisant varier la fréquence et la régularité de collecte de données. Pour la fréquence, nous avons testé un échantillonnage quotidien, un échantillonnage à une fréquence moyenne d'un jour sur deux, un jour par semaine, ou un jour par quinzaine. Hormis pour l'échantillonnage quotidien, deux niveaux de régularité ont été testés pour chaque fréquence. Pour l'échantillonnage réalisé en moyenne tous les deux jours, nous avons découpé la saison de reproduction en paires de jours consécutifs. L'échantillonnage régulier était réalisé en choisissant systématiquement les jours pairs ou systématiquement les jours impairs. L'échantillonnage irrégulier était réalisé en choisissant aléatoirement le premier ou le deuxième jour de chaque paire. Pour l'échantillonnage hebdomadaire et par quinzaine, l'échantillonnage régulier était réalisé en échantillonnant systématiquement le même jour de la semaine ou de la quinzaine, alors que l'échantillonnage irrégulier correspondait au choix aléatoire d'un jour différent pour chaque semaine ou chaque quinzaine. On a donc sept plans d'échantillonnages : 1) quotidien, 2) tous les deux jours et régulier, 3) un jour sur deux et irrégulier, 4) hebdomadaire et régulier, 5) hebdomadaire et irrégulier, 6) par quinzaine et régulier, 7) par quinzaine et irrégulier.es cours d’eau pour évaluer l’écart d’estimation entre les deux méthodes.
Ainsi, nous avons simulé un jeu de données complet avec le modèle individu-centré utilisé dans la procédure ABC. Nous avons ensuite sous-échantillonné ce jeu de données pour simuler des plans d'échantillonnage dégradés, et fait tourner le modèle ABC sur les données issues des plans dégradés. Pour le plan complet, un seul scénario est possible. Pour le plan régulier avec échantillonnage un jour sur deux, deux scénarios sont possibles (jours pairs ou impairs) et ont été réalisés. Pour les plans réguliers avec un échantillon par semaine ou par quinzaine, sept scénarios sont possibles (un par jour de la semaine) et ont été réalisés. Pour les plans irréguliers avec un échantillonnage par semaine ou par quinzaine, de nombreux scénarios sont possibles mais seuls sept ont été réalisés (sélectionnés aléatoirement). Pour chacun des 38 scénarios d'échantillonnage correspondant aux sept plans d'échantillonnage, les données générées ont alimenté le modèle ABC pour donner une estimation d'effectif (valeur estimée et incertitude). Cette estimation d’effectif a été comparée pour les 37 plans d’échantillonnage dégradés à l’estimation réalisée avec un plan d’échantillonnage complet (à défaut de connaître la population réelle).
Les résultats de ces simulations de plans d’échantillonnage (figure 3) indiquent que plus l’échantillonnage est dégradé, plus l’estimation est variable et s’éloigne de celle de l’échantillonnage complet. Alors que l’échantillonnage régulier tous les deux jours donne une estimation entre – 12 % et + 5 % quand celui-ci est comparé aux données complètes, et que l’échantillonnage irrégulier un jour sur deux donne une estimation entre – 6 % et + 13 %, les échantillonnages moins intensifs sont eux plus variables, allant jusqu’à – 55 % et + 42 % pour l’échantillonnage irrégulier une fois toutes les deux semaines. Ces résultats indiquent donc qu’aller sur le terrain un jour sur deux semble être un bon compromis entre justesse de l’estimation et coût. Il est intéressant de remarquer que la régularité de l’échantillonnage n’est visiblement pas un paramètre important, ce qui suggère une facilité d’adaptation du protocole à ce niveau.
Figure 3. Comparaison de l’abondance de reproducteurs estimée par le modèle ABC pour les différents échantillonnages testés, chacun étant répété entre 2 et 7 fois.
Perspectives d’amélioration des performances du modèle
Un premier axe d’amélioration du modèle est l’obtention, sur différents sites, de valeurs de paramètres, afin de comparer celles-ci aux premières estimations réalisées sur le site d’étude de la Nive. En cas de différences notables, il sera alors nécessaire de prendre en compte cette variabilité, et éventuellement de comprendre les facteurs expliquant celle-ci. On peut notamment penser à une variabilité de comportement selon les sites, liée ou non à des variables environnementales. Le cas échéant, maximiser l’hétérogénéité entre les sites suivis permettra de mieux comprendre les mécanismes engendrant les différences de comportement observées. En parallèle de ces estimations de paramètres, la mise en œuvre de protocoles de CMR sur les sites suivis permettra de tester les performances du modèle, comme cela a été réalisé pour la Nive.
Un second axe d’amélioration est la modification du modèle selon les besoins des gestionnaires. En effet, s’il est nécessaire d’obtenir un modèle donnant des estimations suffisamment justes et précises, le protocole réalisé en routine (après la calibration sur d’autres sites précédemment citée) doit demeurer simple et en accord avec les contraintes des structures. Le modèle pourrait notamment s’appuyer sur un comptage du nombre de nids journaliers indépendamment de l’activité observée, et ce sans prise en compte de la présence ou non du nid lors d’un précédent comptage (pour éviter de devoir répertorier les nids déjà existants). Le modèle devra alors être ajusté en conséquence. Une meilleure définition des paramètres par rapport au site pourra par ailleurs améliorer les estimations finales et éventuellement permettre un suivi moins intensif qu’un jour sur deux (meilleur compromis avec le modèle actuel entre justesse et limitation des coûts). En plus de limiter les coûts de suivi sur un site donné, l’intérêt serait également de pouvoir suivre plus de sites au cours d’une même saison. Le modèle ici présenté, bien que déjà fonctionnel, se veut donc évolutif. Cette flexibilité est permise par la possibilité que l'utilisateur change les paramètres dans l'application, et par le partage libre (licence Creative Commons CC BY-NC-SA
Conclusion
Le modèle mis en œuvre donne en l’état des estimations d’effectifs de lamproie marine cohérentes avec le modèle de capture-marquage-recapture, bien que significativement plus faibles pour le modèle ABC. Les paramètres influençant le plus l’estimation ont pu être identifiés et seront à déterminer en priorité sur d’autres sites d’étude que celui ayant servi à mettre en œuvre la version actuelle du modèle. L’application développée permet d’utiliser facilement le modèle et de l’adapter à d’autres sites où le comportement des individus serait différent. En plus de préciser les paramètres utilisés, le modèle pourra être modifié afin de correspondre au mieux aux besoins des structures de gestion, en permettant notamment de limiter le temps passé sur le terrain. À terme, cet outil pourrait être utilisé en routine sur plusieurs sites afin de compléter les suivis actuels réalisés notamment au niveau des stations de comptage.
___________________________
Photo d’entête : © INRAE-GLISE
Notes
- 1. Qualifie les espèces animales marines qui remontent les rivières pour se reproduire en eau douce.
- 2. Un modèle bayésien de type ABC permet d'approximer la distribution a posteriori des paramètres d'un modèle en utilisant des méthodes d'approximation basées sur des comparaisons entre données simulées et données observées.
- 3. Test statistique non paramétrique qui permet de tester l'hypothèse selon laquelle les distributions de deux groupes de données sont proches.
- 4. Langage de programmation open-source largement utilisé pour l'analyse statistique et la manipulation de données.
- 5. La licence CC-by-NC-SA 4.0 permet toute exploitation de l’œuvre (partager, copier, reproduire, distribuer, communiquer, réutiliser, adapter) par tous moyens, sous tous formats. Toutes les exploitations de l’œuvre ou des œuvres dérivées, sauf à des fins commerciales, sont possibles
Références
- Beaumont, M. A. (2010). Approximate Bayesian Computation in evolution and ecology. Annual Review of Ecology, Evolution, and Systematics, 41(1), 379406. https://doi.org/10.1146/annurev-ecolsys-102209-144621
- Chang, W., Cheng, J., Allaire, J., Xie, Y., & MvPherson, J. (2015). Package ‘shiny’ (1.7.2) [Logiciel]. http://shiny.rstudio.com/
- Csilléry, K., Blum, M. G. B., Gaggiotti, O. E., & François, O. (2010). Approximate Bayesian Computation (ABC) in practice. Trends in Ecology & Evolution, 25(7), 410418. doi:10.1016/j.tree.2010.04.001
- Dhamelincourt, M., Buoro, M., Rives, J., Sebihi, S., & Tentelier, C. (2021a). Individual and group characteristics affecting nest building in sea lamprey (Petromyzon marinus L. 1758). Journal of Fish Biology, 98(2), 557565. doi:10.1111/jfb.14601
- Dhamelincourt, M., Tentelier, C., & Elosegi, A. (2023). ABC model for estimating sea lamprey local population size using a simple nest count during the spawning season. Knowledge & Management of Aquatic Ecosystems, 424, 5. https://doi.org/10.1051/kmae/2023002
- Funk, W. C., Almeida-Reinoso, D., Nogales-Sornosa, F., & Bustamante, M. R. (2003). Monitoring population trends of Eleutherodactylus frogs. Journal of Herpetology, 37(2), 245256. doi:10.1670/0022-1511(2003)037
- Johnston, C. A., & Windels, S. K. (2015). Using beaver works to estimate colony activity in boreal landscapes. The Journal of Wildlife Management, 79(7), 10721080. doi:10.1002/jwmg.927
- Kruschke, J. K. (2010). What to believe: Bayesian methods for data analysis. Trends in Cognitive Sciences, 14(7), 293300. doi:10.1016/j.tics.2010.05.001
- Lenormand, M., Jabot, F., & Deffuant, G. (2013). Adaptive approximate Bayesian computation for complex models. Computational Statistics, 28(6), 27772796. doi:10.1007/s00180-013-0428-3
- McClintock, B. T., & White, G. C. (2009). A less field-intensive robust design for estimating demographic parameters with Mark-resight data. Ecology, 90(2), 313-320. doi:10.1890/08-0973.1
- Morgan, B. J. T., North, P. M., Ralph, C. J., & Scott, J. M. (1983). Estimating numbers of terrestrial birds. Biometrics, 39(4), 1123. doi:10.2307/2531357
- Schwarz, C. J., & Seber, G. A. F. (1999). Estimating animal abundance: Review III. Statistical Science, 14(4), 427456. https://www.jstor.org/stable/2676809
Résumé
L’estimation des effectifs d’une population nécessite de prendre en compte la biologie de l’espèce mais aussi des contraintes logistiques comme le coût humain et matériel. Les méthodes classiques basées sur des comptages individuels fournissent des estimations précises mais sont en général lourdes à mettre en œuvre. En parallèle, les méthodes basées sur le recensement d’indices de présence ne permettent en général pas une estimation absolue de la population. Une manière de résoudre cette limite est de connaître la relation entre les indices de présence et l’effectif de population. Dans ce papier, nous présentons un modèle simulant une saison de reproduction de lamproie marine et liant le comportement individuel et le nombre de nids construits pour estimer le nombre de reproducteurs. Les données d’entrée du modèle sont un nombre de nids journalier compté sur le terrain. Ce modèle, paramétré à partir d'observations comportementales sur le terrain, donne des estimations de population réalistes, le rendant fonctionnel en l’état. Il est par ailleurs implémenté dans une application en ligne facile à utiliser et modifier et ne nécessitant pas de connaissance de codage. Nous discutons les possibilités d’amélioration et d’adaptation aux contraintes rencontrées sur le terrain afin de rendre ce modèle adapté aux besoins de gestion.
Pas de document complémentaire pour cet article
Statistiques de l'article
 Vues: 7534
Vues: 7534
Téléchargements
 PDF: 220
PDF: 220
 XML: 61
XML: 61