

SalmoGlob – Un nouveau modèle pour l’évaluation des stocks de saumon atlantique à l’échelle de l’Atlantique Nord
Évaluer l'état des populations de poissons sauvages est essentiel pour guider une gestion efficace dans un contexte de fortes pressions sur la ressource. Dans le cadre du projet SalmoGlob, les chercheurs ont développé un modèle innovant pour évaluer les stocks de saumon atlantique dans l’Atlantique Nord. Ce modèle intègre des données depuis 1971, permettant d’analyser la dynamique des populations à différentes échelles et stades de vie. Adopté par le Conseil international pour l’exploration de la Mer en 2024, il améliore significativement la qualité des évaluations scientifiques et des scénarios de gestion face aux changements globaux.
Introduction
Évaluer le statut des populations de poissons sauvages est un prérequis pour pouvoir éclairer une gestion efficace dans un contexte de fortes pressions anthropiques. Cela consiste à estimer l’abondance des populations ainsi que son évolution dans le temps et dans l’espace et à la comparer à des points de référence. Il s’agit aussi d’appréhender les changements potentiels dans la structure démographique des populations, par exemple dans la structure d’âge, une moindre diversité de la structure d’âge étant généralement un signe de mauvaise santé des populations.
La démarche d’évaluation des stocks halieutiques s’est historiquement développée avec comme objectif de quantifier l’impact de l’exploitation par la pêche sur l’abondance et sur la structure démographique des populations, la pêche étant perçue comme la source de pression principale exercée sur les populations. Dans le cadre de la transition vers une approche écosystémique de la gestion des pêches (Garcia, 2003), les recommandations internationales encouragent à faire évoluer l’évaluation des stocks vers une approche plus holistique du fonctionnement des populations en interaction avec les composantes de l’écosystème. Cela consiste notamment à développer une nouvelle génération de modèles permettant de quantifier l’influence de multiples facteurs de pression sur les populations, au-delà de la seule exploitation par la pêche, comme la dégradation des habitats ou les impacts directs et indirects du changement climatique.
Cela soulève de nombreux défis, en particulier pour les poissons migrateurs amphihalins comme le saumon atlantique (Salmo salar). Répartie dans l'océan Atlantique Nord et la mer Baltique, cette espèce partage son cycle de vie entre un habitat de reproduction et de croissance des juvéniles en eau douce et un habitat de croissance et de maturation en mer. Les saumons qui se reproduisent dans les cours d’eau des côtes Atlantique de l’Amérique du Nord et de l’Europe entreprennent de longues migrations marines vers des zones de nourrissage communes à plusieurs populations, où ils sont exposés à des conditions environnementales océaniques communes ainsi qu’à des pêcheries communes. Après ce séjour marin, ils rejoignent leur cours d’eau natal pour se reproduire.
Une telle dépendance à plusieurs écosystèmes rend cette espèce sensible à de multiples facteurs de pression à différents stades de vie et à des échelles spatiales emboîtées, allant de facteurs très locaux agissant spécifiquement sur une population (par exemple, la modification de l’habitat continental ou de l’exploitation par la pêche récréative dans les cours d’eau) à des facteurs pouvant agir simultanément sur un grand nombre de populations (par exemple, la modification de l’écosystème marin ou l’exploitation en mer sur des mélanges de populations) (Olmos et al., 2020). Cela génère de fortes dépendances spatiales entre les populations, de nature écologique mais aussi en termes de gestion, qui appellent la mise en place de modèles d’évaluation prenant en compte des emboîtements d’échelles spatiales allant du local jusqu’à l’ensemble du bassin de l’Atlantique Nord (Olmos et al., 2019 ; Bull et al., 2022).
La mise en place d’outils d’évaluation est d’autant plus critique que l’abondance de nombreuses populations de saumons a considérablement diminué, d’un facteur 4 entre le début des années 1970 et aujourd’hui (Limburg et Waldman, 2009 ; ICES, 2024)
La surexploitation historique par la pêche est l’une des pressions responsables de ce déclin dont les causes sont multifactorielles. Les stocks de saumon ont été intensément exploités entre les années 1960 et le début des années 1990, avec des captures totales dans l’Atlantique Nord qui ont avoisiné les douze mille tonnes par an au début des années 1970 (Chaput et al., 2012). Des pêcheries importantes opérant sur des mélanges de populations se sont développées au niveau des zones communes de nourrissage situées au large des îles Féroé et du Groenland, avant d’être fermées en 1992 (Féroé) et 1998 (Groenland ; seule la pêcherie de subsistance reste autorisée depuis 1998). D’autres pêcheries opèrent dans des zones plus côtières où les saumons transitent avant leur entrée dans les cours d’eau pour la reproduction, ou bien en zones estuariennes ou dans les cours d’eau juste avant la reproduction. La fermeture progressive des principales pêcheries de saumons n’a pas suffi à enrayer le déclin des populations, dont la persistance est attribuée à une baisse du taux de retour des individus après leur séjour marin (un proxy de la survie en mer) en réponse à des changements de l’écosystème marin à grande échelle, en lien avec la diminution de la qualité et de la quantité des proies disponibles pour le saumon pendant la phase marine (ICES, 2017 ; Olmos et al., 2020 ; ICES, 2023a).
Dans ce contexte, l’urgence de mettre en place des mesures de gestion au niveau international a conduit à la création, en 1982, de l’Organisation pour la conservation du saumon de l’Atlantique Nord (NASCO/OCSAN
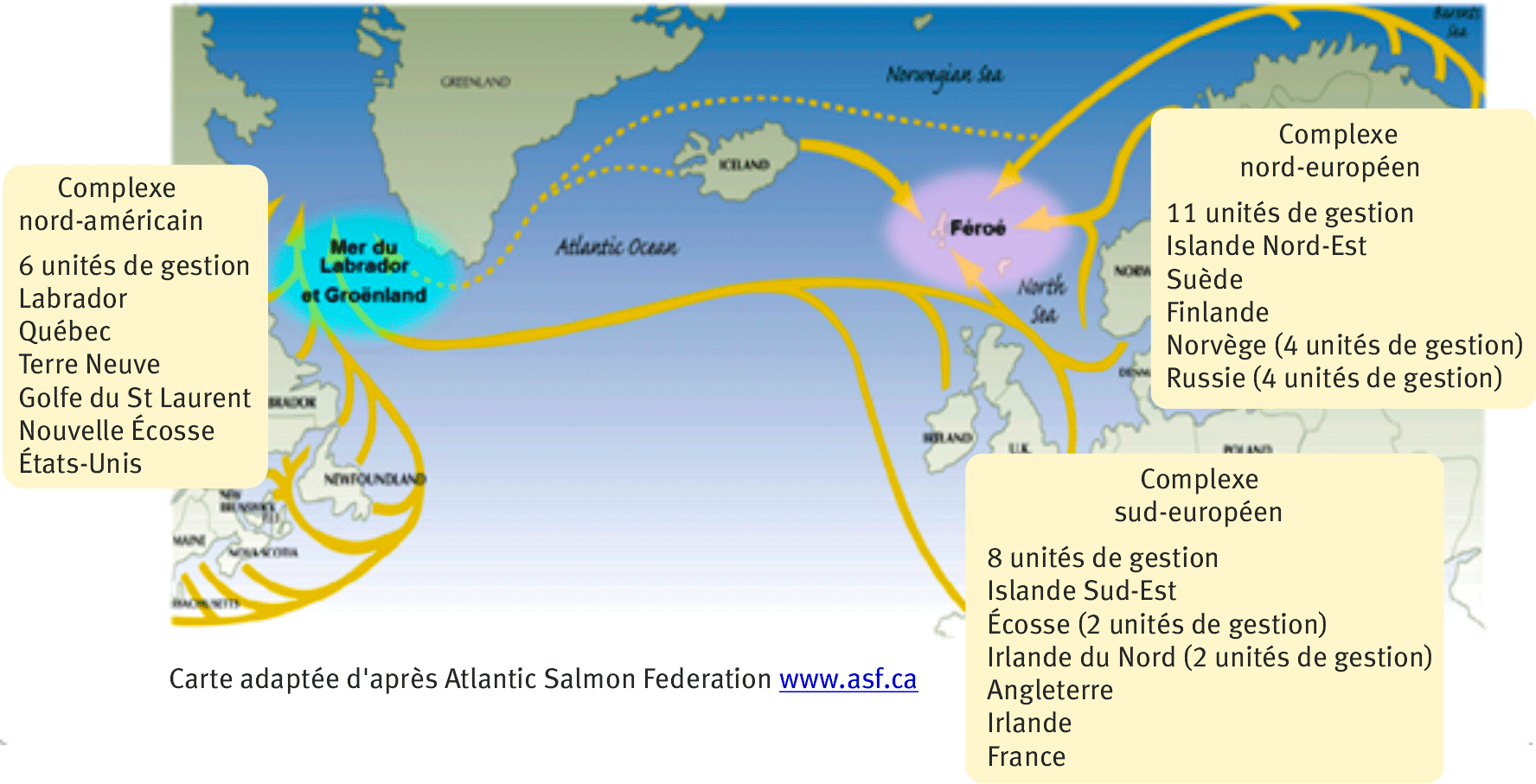
Des modèles ont été élaborés par le WGNAS afin d’évaluer l’état des populations à l’échelle des unités de gestion (UG) régionales ou nationales de tout le bassin océanique de l'Atlantique Nord (Crozier et al., 2004 ; Potter et al., 2004). Ces unités de gestion (vingt-cinq à l’heure actuelle ; figure 1) sont regroupées en trois complexes de populations continentaux : le complexe sud-européen, nord-européen et nord-américain (Chaput et al., 2012). Ces modèles permettent d’estimer des séries à long terme (du début des années 1970 à aujourd’hui) d’abondance en mer avant toute exploitation par la pêche et de prévoir les abondances des saumons adultes au retour dans les cours d’eau trois années en avance. Ces projections tiennent compte des variations de la productivité des populations et des diverses pêcheries séquentielles avant la reproduction. Ces modèles sont incorporés dans un cadre d'analyse de risques pour évaluer l’abondance des retours de reproducteurs vis-à-vis des limites de conservation (points de référence en dessous desquels l’abondance ne doit pas passer pour garantir la conservation des populations) pour chaque UG et au niveau plus agrégé des complexes continentaux. Ils permettent de quantifier les conséquences des variations de la productivité marine et des pêcheries opérant sur des mélanges de populations au Groenland et sur l’abondance des retours aux îles Féroé, et ainsi de porter un avis sur l’opportunité de rouvrir les pêcheries commerciales en mer.
Figure 1. Les vingt-cinq unités de gestion (UG) pour le saumon atlantique de l’Atlantique Nord considérées par le groupe de travail sur le saumon atlantique (WGNAS).
Représenter la dynamique de toutes les unités de gestion de l’Atlantique Nord dans un cadre harmonisé
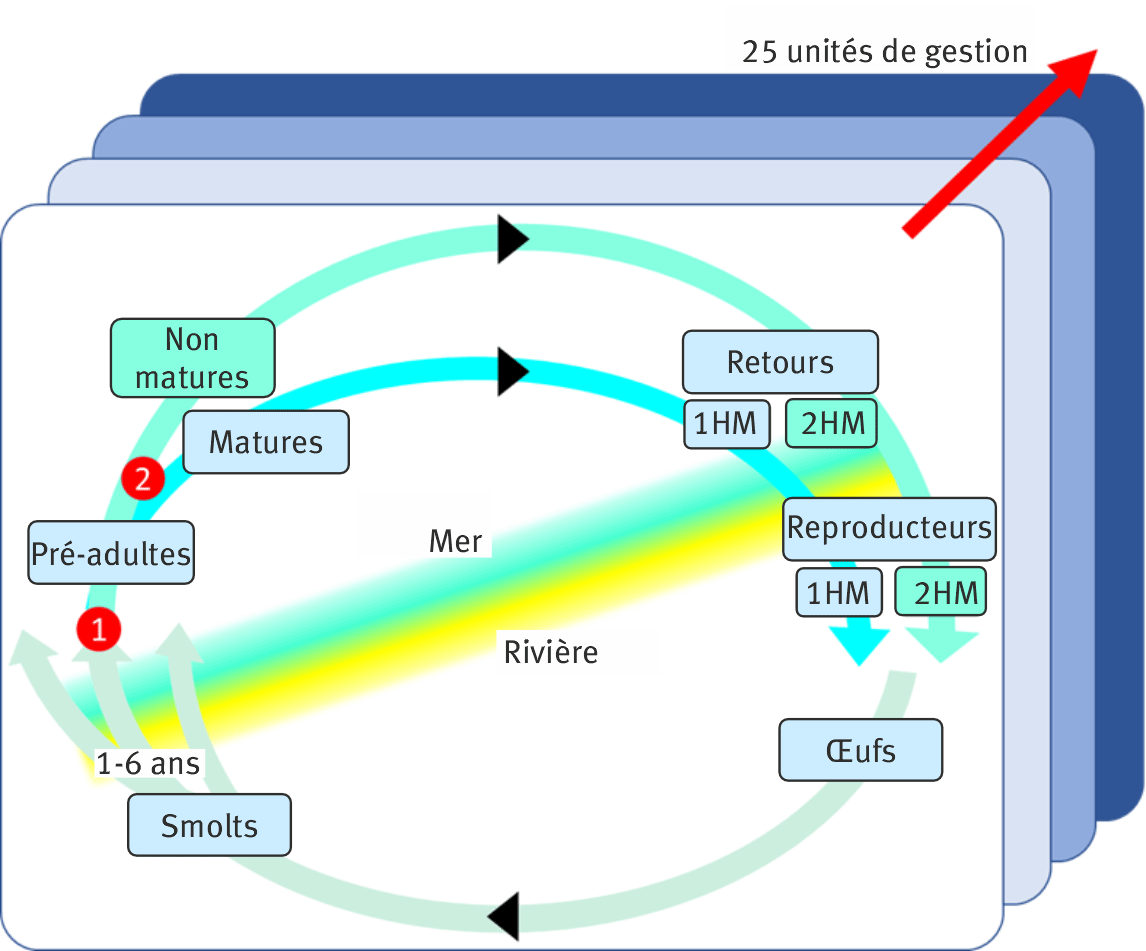
Au cœur du nouveau cadre de modélisation, nous avons développé un modèle de cycle de vie structuré en âges et en stades de vie qui représente explicitement la dynamique des abondances entre le stade œuf et le stade adulte reproducteur de retour dans sa rivière natale pour la reproduction après le séjour en mer (figure 2). La dynamique des populations des vingt-cinq UG de l’Atlantique Nord est représentée simultanément dans le même modèle (Olmos et al., 2019 ; Rivot et al., 2024). Le modèle est construit dans un cadre hiérarchique bayésien (Parent et Rivot, 2012 ; Gimenez et Peyrard, 2022), qui permet d’ajuster la dynamique aux observations disponibles dans toutes les unités de gestion entre 1971 et aujourd’hui, en intégrant plusieurs sources de données de nature hétérogène (encadré 1).
Le nouveau modèle permet d’analyser la dynamique des populations, de réaliser l’évaluation des stocks et d’évaluer des scénarios de gestion simultanément pour l’ensemble des vingt-cinq UG de l’Atlantique Nord, dans un cadre cohérent et unifié. Cela représente un progrès majeur en termes de réalisme biologique et de compa-rabilité des estimations par rapport aux modèles historiquement utilisés par le WGNAS, qui s’appuyaient sur trois modèles indépendants et différents pour les complexes nord-américain, sud-européen et nord-européen. Cette unification rationalise et simplifie la chaine de traitement des données et harmonise la structure démographique et la représentation des stades de vie entre toutes les UG d’Amérique du Nord et d’Europe (figure ). En particulier, dans le modèle utilisé historiquement pour l’Amérique du Nord, seule l’abondance des poissons passant deux hivers en mer avant la reproduction (2HM) était considérée. Cela revient à ignorer les poissons passant un hiver en mer avant leur retour (1HM) dans la démographie et la dynamique de population, alors que cette composante peut représenter une part non négligeable de l’abondance dans certaines UG (ICES, 2024). Dorénavant, le nouveau modèle de cycle de vie intègre la dynamique des deux classes d’âge de mer 1HM et 2HM pour toutes les UG.
Figure 2. Modèle de cycle de vie pour les vingt-cinq unités de gestion (UG) du bassin de l’Atlantique Nord.
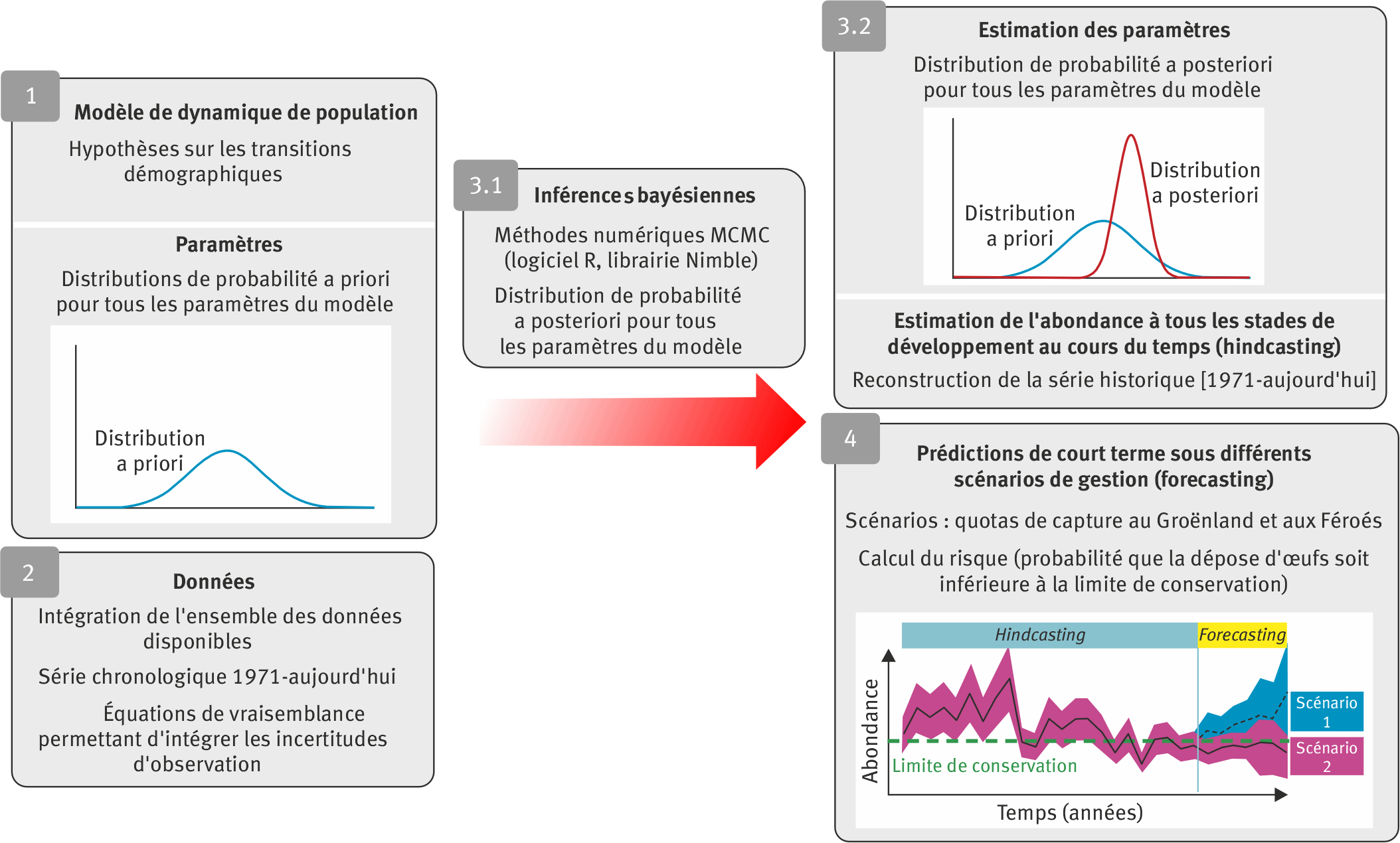
Étape 1. Les hypothèses de modélisation sont posées sur la structure du modèle de dynamique de population (définition des stades de développement, paramétrisation des transitions démographiques entre les stades, liens potentiels entre les différentes unités de gestion). Des hypothèses supplémentaires sont faites sur la distribution de probabilité a priori des paramètres du modèle. Dans le cas présent, des distributions a priori peu informatives ont été choisies.
Étape 2. Les données sont intégrées dans le modèle, au travers d’équations de vraisemblance qui permettent de considérer les erreurs d’observation.
Étape 3. L’inférence bayesienne (étape 3.1) consiste à combiner les hypothèses a priori avec les données pour obtenir (étape 3.2) les distributions a posteriori de tous les paramètres du modèle ainsi que des abondances dans tous les stades de développement au cours du temps (« hindcasting »). Dans la pratique, cette étape fait appel à des simulations numériques de type Monte Carlo par Chaine de Markov (MCMC) réalisées sous le logiciel R grâce à la librairie nimble (https://r-nimble.org).
Étape 4. Le modèle est utilisé pour réaliser des prédictions à court terme (un à quatre ans dans le cas présent) sous différents scénarios. Le cadre bayesien permet de propager les incertitudes dans les prédictions. Les risques associés à chaque scenario sont calculés comme les probabilités que les abondances d’œufs soient inférieures aux limites de conservation définies pour chaque unité de gestion.
Mieux comprendre la réponse des populations aux changements globaux
La formulation de la dynamique de population de l’ensemble des UG de l’Atlantique Nord dans un cadre harmonisé permet de mieux comprendre les mécanismes de la réponse des populations de saumon à de multiples facteurs de forçage agissant sur les populations à des échelles spatiales différentes (exploitation et modification de l’environnement, notamment marin).
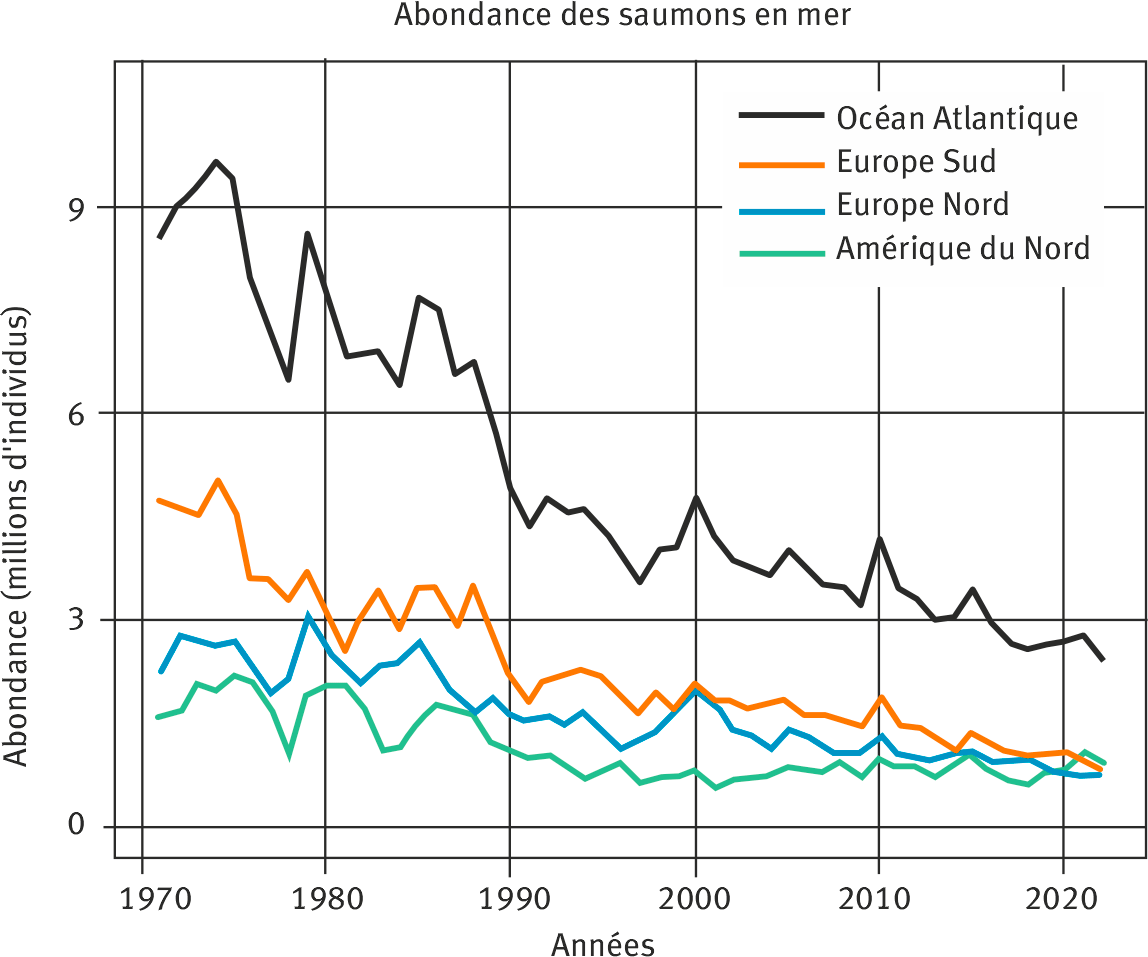
Les estimations issues du modèle confirment un fort déclin de l’abondance des saumons en mer, d’un facteur 3 à 4 depuis le début des années 1970 (figure 3).
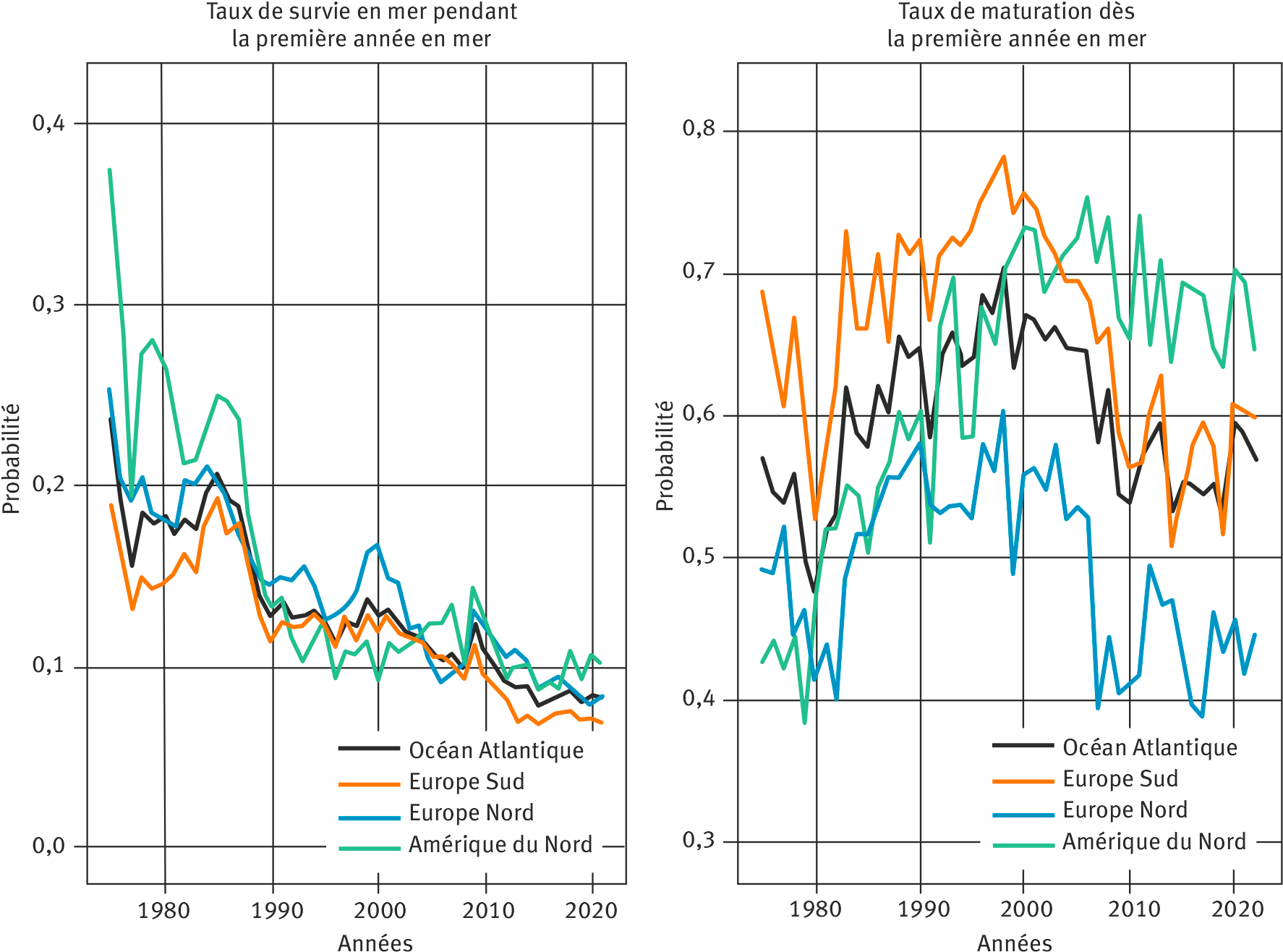
Le modèle permet d’estimer un ensemble de paramètres de la démographie et de la dynamique de population simultanément pour l’ensemble des UG de l’Atlantique Nord, et notamment : i) l’abondance des saumons en mer avant toute mortalité par pêche ; ii) le taux de survie des saumons pendant la première année en mer ; iii) le taux de maturation à la fin de la première année en mer (figure 4). Les résultats mettent en évidence la forte cohérence spatiale dans les variations temporelles de la survie des post-smolts et dans la proportion de poissons qui deviennent matures dès la fin de la première année en mer entre différentes UG qui partagent des routes de migration marines communes. À l’échelle du bassin de l’Atlantique Nord, le taux de survie pendant la première année en mer a chuté de près de 70 % depuis le début des années 1970. La probabilité de déclencher la maturation sexuelle dès la fin de la première année passée en mer a augmenté jusque dans les années 1990 (synonyme d’une baisse de la proportion de poissons âgés de plusieurs hivers de mer dans les retours) avant d’atteindre un plateau voire de décroitre sur la période récente (surtout pour les poissons européens). Le signal commun entre les différentes UG explique plus de 40 % de la variabilité temporelle totale, et la covariation spatiale entre les UG augmente avec la proximité géographique (Olmos et al., 2019 ; Rivot et al., 2024).
Cette synchronie dans les variations temporelles suggère une réponse des populations à un signal environnemental agissant à large échelle. En s’appuyant sur ce modèle, Olmos et al. (2020) ont démontré que les variations de la survie étaient corrélées avec des proxys des conditions environnementales et des ressources trophiques disponibles pour le saumon dans des fenêtres spatio-temporelles correspondant aux habitats marins occupés par de multiples populations pendant la fin de l’été et le début de l’automne, en Mer de Norvège pour les populations européennes, et dans la mer du Labrador au sud et à l’ouest du Groenland pour les populations nord-américaines. Les auteurs ont démontré que les variations dans la survie en mer étaient négativement corrélées aux variations de la température de surface de la mer et positivement corrélées avec les variations de la productivité primaire estimées à partir de données satellite pour ces fenêtres spatio-temporelles. Ces résultats renforcent l’hypothèse d’une réponse globale des populations aux changements dans l’écosystème marin. Ils soulignent le potentiel de ce nouveau modèle pour l’intégration de variables environnementales afin de tester des hypothèses écologiques sur l’influence des changements globaux et d’augmenter sa capacité explicative et prédictive.
Évaluer des scénarios dans un cadre probabiliste et une approche de précaution
Le modèle de cycle de vie offre un cadre naturel pour réaliser des projections de la dynamique de population sous divers scénarios pouvant faire intervenir des pressions (environnement et exploitation par la pêche) qui affectent les populations à plusieurs échelles spatiales, allant de certaines UG spécifiques jusqu’à des groupes de plusieurs UG à une échelle continentale. Le même modèle de cycle de vie est utilisé à la fois pour la phase d’ajustement aux séries de données historiques (hindcasting) et de projection de quelques années dans le futur après la dernière année de données disponibles (forecasting). L’intégration des vingt-cinq UG au sein d’un même modèle permet de réaliser des projections de la dynamique de population et des retours pour toutes les UG simultanément.
Toutes les propriétés statistiques du modèle sont automatiquement transférées dans la phase de projection et dans les analyses de risque. La covariation spatiale des séries temporelles du taux de survie en mer et du taux de maturation dès la fin de la première année en mer est prise en compte dans les projections. Toutes les sources d’incertitude liées à la stochasticité temporelle des principaux taux de transition démographiques et à l’estimation des paramètres sont propagées dans les prédictions pour réaliser des analyses de risque dans un cadre probabiliste.
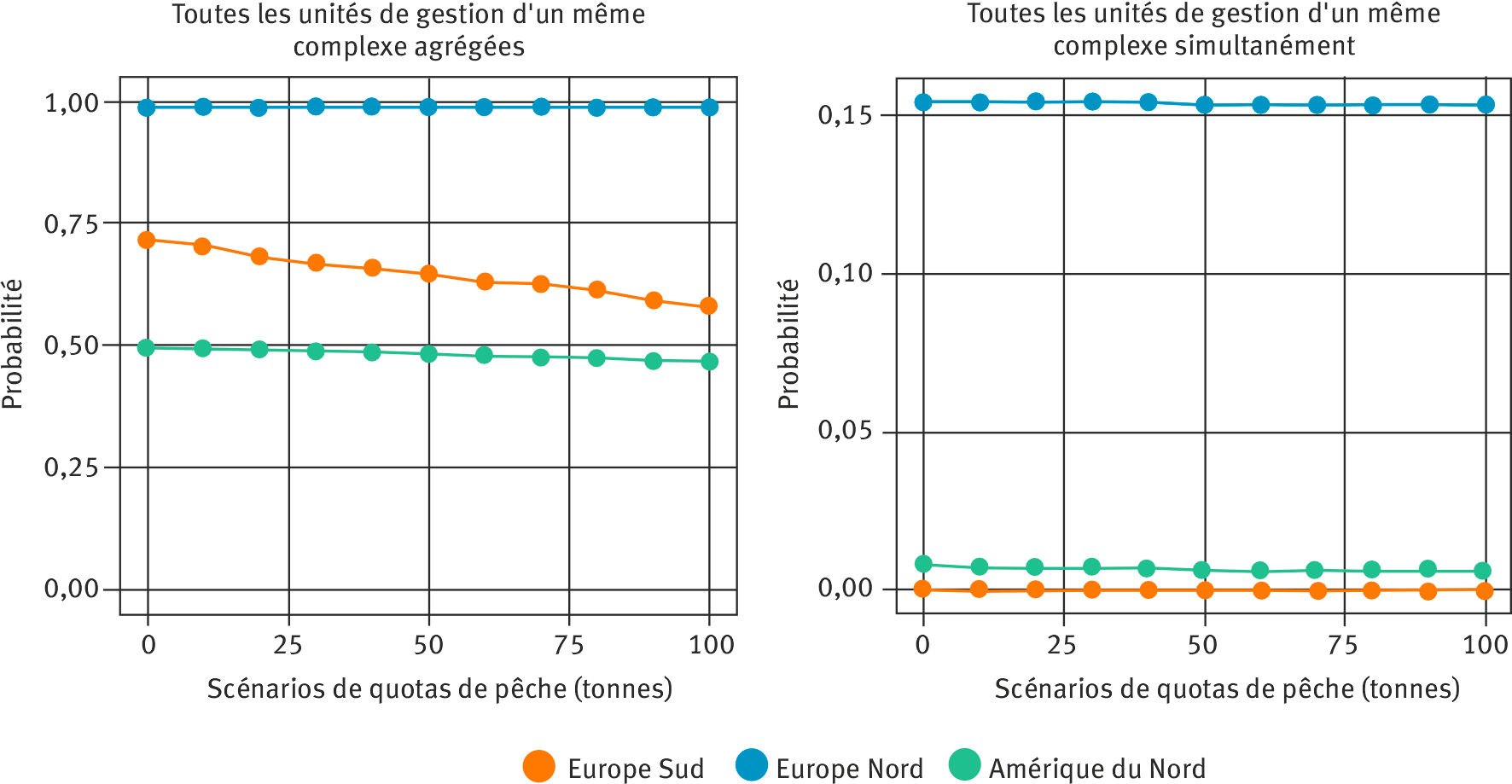
Par exemple, il est possible d’évaluer la probabilité que l’abondance des retours dans chaque UG atteigne les limites de conservation selon différents scénarios de quotas de pêche au large du Groenland (pêche qui impacte les UG nord-américaines et européennes), ainsi que la probabilité que toutes les UG d’un même complexe de populations atteignent simultanément leur limite de conservation (figure 5). Actuellement, ces probabilités sont très faibles (proche de 0) surtout pour les complexes de populations de l’Europe du Sud et de l’Amérique du Nord, ce qui conduit le WGNAS à préconiser de ne pas rouvrir la pêche commerciale au Groenland (ICES, 2024). Le modèle pourrait aussi être utilisé pour évaluer l’impact d’autres pêcheries, ou pour évaluer des mesures de conservation spécifiques pour les différentes classes d’âge.
Figure 5. Probabilité que l’abondance des retours de saumons adultes prédite pour l’année 2026 (projection à trois ans après dernière année évaluée) atteigne les limites de conservation (LC), en fonction de différents scenarios de quotas de captures au Groenland (en tonnes de poissons).
Un outil opérationnel conçu dans une logique de science ouverte
Afin de faciliter son appropriation par la communauté, une suite de programmes a aussi été conçue dans une logique de science ouverte et intégrée dans le portail GitHub du CIEM (à ce jour encore en accès restreint au WGNAS). Cette suite de programmes est développée dans le langage R et mobilise le package nimble pour l'inférence bayésienne. Elle facilite l’utilisation du modèle dans le contexte de temps très contraint de la réunion annuelle du WGNAS. En outre, une base de données et une application disponible sur le web
Conclusion
Ce nouveau cadre de modélisation a fait l’objet d’une procédure d’évaluation et de validation par les instances du CIEM (procédure de benchmarking – ICES, 2023b) qui s’est concrétisée par son adoption pour l’évaluation des stocks et les avis rendus à l’OCSAN depuis 2024 (ICES, 2023b ; ICES, 2024). Les produits du projet SalmoGlob ont d’ores-et-déjà profondément modifié les travaux du groupe de travail WGNAS et le potentiel évolutif du modèle permet d’envisager une démarche d’amélioration progressive dans la durée.
En plus des avantages proposés dans la version actuelle du modèle et utilisé pour l’avis émis en 2024, ce nou-veau cadre de modélisation se révèle flexible et évolutif. Le fait d’utiliser un modèle de cycle de vie structuré en âges et en stades a enclenché une réappropriation du cadre de modélisation utilisé pour l’évaluation et l’avis par la communauté des biologistes et des personnes impliquées dans la récolte des données. Cette dynamique vertueuse a déjà permis d’intégrer dans le modèle de nouvelles données et connaissances jusqu’à présent sous-utilisées. Par exemple, les informations disponibles sur le changement de la fécondité moyenne des saumons observé dans la dernière décennie (en lien avec la baisse de la taille moyenne à maturité) ont pu être prises en compte. La flexibilité du cadre de modélisation a aussi encouragé l’intégration de nouvelles données génétiques pour informer la répartition des captures au Groenland entre les différentes UG. Cela apporte un degré de réalisme supplémentaire en permettant de mieux représenter la façon dont les pêcheries opérant sur des mélanges de populations influencent les différentes UG.
Ainsi, ce modèle flexible et évolutif offre une agilité accrue pour intégrer de nouvelles connaissances et adapter les mesures de gestion dans un contexte de changement environnemental rapide.
Rivot, E., Olmos, M., Nevoux, M., Prévost, E., Buoro, M., & Beaulaton, L. (2020). SalmoGlob - ToolBoxWGNAS. Développement d’un nouveau modèle pour l’évaluation des stocks de saumon atlantique à l’échelle de l’Atlantique Nord. Rapport de recherche. OFB-Inrae-Institut Agro-Agrocampus Ouest-UPPA. 75 p. https://hal.science/hal-03138237
Remerciements
Ce travail a été rendu possible grâce à l’investissement de nombreuses personnes qui collectent et compilent les données utilisées par le groupe de travail WGNAS en Amérique du Nord et en Europe (et notamment pour la France). Le projet a été financé par la convention de coopération du Pôle OFB, INRAE, l’Institut Agro, UPPA Gestion des Poissons Migrateurs Amphihalins dans leur Environnement. Le projet a aussi reçu le soutien de l’Université de Rennes 1 et du fonds de développement régional de l’Europe au travers du programme Interreg France-Manche-Angleterre, projet SAMARCH Salmonid Management Round the Channel.
_____________________________________
Photo d’entête : © Guillaume Evanno / INRAE
Notes
- 1. ICES : International Council for the Exploration of the Sea. En français : CIEM : Conseil International pour l’Exploration de la Mer (CIEM).
- 2. IUCN : Union internationale pour la conservation de la nature.
- 3. NASCO : North Atlantic Salmon Conservation Organization ; OCSAN : Organisation pour la conservation du saumon de l’Atlantique Nord.
- 4. WGNAS : Working Group on North Atlantic Salmon.
- 5. http://sirs.agrocampus-ouest.fr/discardless_app/WGNAS-ToolBox/
Références
- Bull, C. D., Gregory, S. D., Rivot, E., Sheehan, T. F., Ensing, D., Woodward, G., & Crozier, W. (2022). The likely suspects frame-work: The need for a life cycle approach for managing Atlantic salmon (Salmo salar) stocks across multiple scales. ICES Journal of Marine Science, 79, 1445-1456. doi:10.1093/icesjms/fsac099
- Chaput, G. (2012). Overview of the status of Atlantic salmon (Salmo salar) in the North Atlantic and trends in marine mortality. ICES Journal of Marine Science, 69(9), 1538-1548. doi:10.1093/icesjms/fss013
- Crozier, W. W., Schön, P. J., Chaput, G., Potter, E. C. E., Maoiléidigh, N. Ó., & MacLean, J. C. (2004). Managing Atlantic salmon (Salmo salar L.) in the mixed stock environment: Challenges and considerations. ICES Journal of Marine Science, 61(8), 1344-1358. doi:10.1016/j.icesjms.2004.08.013
- Darwall, W. R. T. (2023). Salmo salar. The IUCN Red List of Threatened Species 2023, e.T19855A67373433. doi:10.2305/IUCN.UK.2023-1.RLTS.T19855A67373433.en
- Garcia, S. (2003). The ecosystem approach to fisheries: Issues, terminology, principles, institutional foundations, implementation and outlook. Food and Agriculture Organisation of the United Nations (FAO).
- Gimenez, O., & Peyrard, N. (2022). Statistical Approaches for Hidden Variables in Ecology. John Wiley & Sons, ISTE.
- Hernvann, P. Y., Lemaire-Patin, R., Guitton, J., Olmos, M., Etienne, M. P., Labouyrie, M., Bezier, L., & Rivot, E. (2024). WGNAS-SalmoGlob ToolBox: a web application for supporting Atlantic salmon stock assessment at the North Atlantic basin scale ICES WGNAS 2024, 11th-21th March 2024, Galway, Ireland. (Working Paper 28).
- ICES. (2017). Report of the Workshop on Potential Impacts of Climate Change on Atlantic Salmon Stock Dynamics (WKCCISAL), 27–28 March 2017, Copenhagen, Denmark. ICES CM 2017/ACOM:39. doi:10.17895/ices.pub.19290416
- ICES. (2023a). The Second ICES/NASCO Workshop on Salmon Mortality at Sea (WKSalmon2; outputs from 2022 meeting). ICES Scientific Reports. 5(36). https://doi.org/10.17895/ices.pub.22560790
- ICES. (2023b). Benchmark Workshop on Atlantic Salmon (Salmo salar) in the North Atlantic (WKBSALMON). ICES Scientific Reports, 5(112). doi:10.17895/ices.pub.24752079.v1
- ICES. (2024). Working Group on North Atlantic Salmon (WGNAS). ICES Scientific Reports, 6(36). doi:10.17895/ices.pub.25730247.v1
- Limburg, K. E., & Waldman, J. R. (2009). Dramatic declines in North Atlantic diadromous fishes. BioScience, 59(11), 955-965. doi:10.1525/bio.2009.59.11.7
- Olmos, M., Massiot-Granier, F., Prévost, E., Chaput, G., Bradbury, I. R., Nevoux, M., & Rivot, E. (2019). Evidence for spatial coherence in time trends of marine life history traits of Atlantic salmon in the North Atlantic. Fish and Fisheries, 20(2), 322-342. doi:10.1111/faf.12345
- Olmos, M., Payne, M. R., Nevoux, M., Prévost, E., Chaput, G., Du Pontavice, H., Guitton, J., Sheehan, T., Mills, K., & Rivot, E. (2020). Spatial synchrony in the response of a long range migratory species (Salmo salar) to climate change in the North Atlantic Ocean. Global Change Biology, 26,1319-1337. doi:10.1111/gcb.14913
- Parent, E., & Rivot, E. (2012). Introduction to hierarchical bayesian modeling for ecological data. Chapman & Hall/CRC.
- Potter, E., Crozier, W., Schön, P., Nicholson, M., Maxwell, D., Prévost, E., Erkinaro, J., Gudbergsson, G., Karlsson, L., Hansen, L., MacLean, J., Maoiléidigh, N. Ó., & Prusov, S. (2004). Estimating and forecasting pre-fishery abundance of Atlantic salmon (Salmo salar L.) in the Northeast Atlantic for the management of mixed-stock fisheries. ICES Journal Of Marine Science, 61(8), 1359-1369. https://doi.org/10.1016/j.icesjms.2004.08.012
- Rivot, E., Lemaire-Patin, R., Olmos, M., Chaput, G., Hernvann, P. Y., & Dubost, G. (2024). A hierarchical Bayesian life cycle model for Atlantic salmon stock assessment and provision of catch advice at the North Atlantic basin scale (Working Paper No. 26). ICES WGNAS 2024, 11–21 March 2024, Galway, Ireland.
Résumé
Le projet SalmoGlob mené au sein de l’unité mixte de recherche DECOD et du pôle MIAME, en partenariat avec le groupe de travail sur le saumon de l’Atlantique Nord du Conseil International pour l’Exploration de la Mer (ICES/CIEM WGNAS), a significativement renforcé les bases scientifiques de l'évaluation des stocks de saumon atlantique (Salmo salar) de l’Atlantique Nord. Un nouveau modèle a été développé pour analyser la dynamique des populations de saumon atlantique de l’ensemble des vingt-cinq unités de gestion (UG) des trois complexes de populations de l’Atlantique Nord : sud-européen, nord-européen et nord-américain. Il intègre plusieurs sources de données hétérogènes disponibles dans toutes les UG depuis 1971. Il peut être étendu pour mieux représenter la démographie et assimiler de nouvelles sources de connaissances et de données. Il représente la dynamique des populations sous la forme d’un modèle structuré en âges et stades de vie, ce qui permet d’intégrer les connaissances disponibles sur la biologie et l’écologie de l’espèce et de représenter l’effet des pressions environnementales à différentes échelles et stades de vie. Le modèle aide à comprendre et à prédire l’effet des changements globaux sur la dynamique et la productivité des populations, et à évaluer des scénarios de gestion de l’exploitation par la pêche dans un cadre probabiliste prenant en compte l’ensemble des sources d’incertitude. Une suite logicielle et une application web ont été développées pour simplifier et renforcer la robustesse de l’ensemble de la chaine de traitement des données jusqu’aux résultats. Ce nouveau modèle représente un saut qualitatif important par rapport aux outils d’évaluation précédents. Il permet d’améliorer la qualité de l’avis scientifique rendu sur l’état des populations pour éclairer la gestion dans le contexte de l’adaptation aux changements globaux. Ce nouveau cadre de modélisation a été officiellement adopté pour l’évaluation internationale du saumon par le CIEM en 2024.
Pas de document complémentaire pour cet article
Statistiques de l'article
 Vues: 7706
Vues: 7706
Téléchargements
 PDF: 234
PDF: 234
 XML: 46
XML: 46